”Hur lång tid tar de olika stadierna i cellcykeln?
hur lång tid tar de olika stadierna i cellcykeln?
Reader Mode
replikering är en av kännetecknen för levande materia. Den uppsättning processer som kallas cellcykeln som genomförs när en cell blir två har varit ett dominerande forskningstema i den molekylära eran med applikationer som sträcker sig långt och brett inklusive studier av sjukdomar som cancer som ibland karakteriseras som en sjukdom i cellcykeln gått fel. Cellcykler är intressanta både för hur de liknar från en celltyp till nästa och för hur de är olika. För att få ämnet i lättnad betraktar vi cellcyklerna i en mängd olika organismer inklusive en modellprokaryot, för däggdjursceller i vävnadsodling och under embryonal utveckling i fruktflugan. Specifikt frågar vi vilka enskilda steg som vidtas för att en cell ska delas upp i två och hur lång tid tar dessa steg?
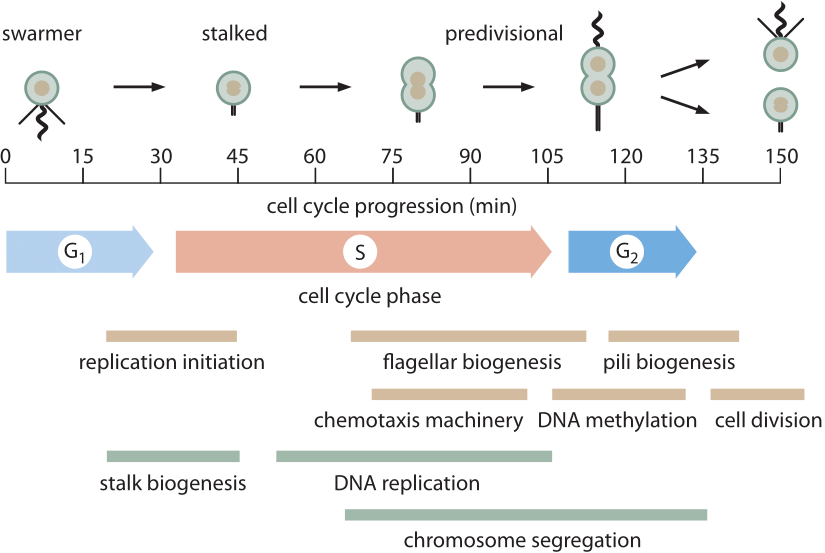
Figur 1: 150 min cellcykel av Caulobacter visas, vilket belyser några av de viktigaste morfologiska och metaboliska händelserna som äger rum under celldelning. M-fas indikeras inte eftersom det i Caulobacter inte finns någon sann mitotisk apparat som monteras som i eukaryoter. Mycket av kromosomsegregering i Caulobacter (och andra bakterier) sker samtidigt med DNA-replikation. De sista stegen i kromosomsegregering och särskilt dekatenering av de två cirkulära kromosomerna sker under G2-fasen. (Anpassad från M. T. Laub et al., Vetenskap 290: 2144, 2000.)
förmodligen är den bäst karakteriserade prokaryota cellcykeln den hos modellorganismen Caulobacter crescentus. En av de tilltalande egenskaperna hos denna bakterie är att den har en asymmetrisk celldelning som gör det möjligt för forskare att binda en av de två avkommorna till ett mikroskopskydd medan den andra dottern driver bort vilket möjliggör ytterligare studier utan hinder. Detta har gett upphov till noggranna skildringar av den 150 minuters cellcykeln i 104921 (bnid) som visas i Figur 1. Huvudkomponenterna i cellcykeln är G1 (första tillväxtfasen, 30 min, bnid 104922), där åtminstone en viss minimal mängd ökning av cellstorleken måste äga rum, S-fas (syntes, 80 min, bnid 104923) där DNA replikeras och G2 (andra tillväxtfasen, 25 min, BNID 104924) där kromosomsegregering utvecklas vilket leder till celldelning (slutfasen varar 15 min). Caulobacter crescentus ger ett intressant exempel på hur vissa organismer främjas till ”modellorganism” – status eftersom de har någon speciell egenskap som gör dem särskilt lämpliga för frågan om intresse. I detta fall går cellcykelprogressionen hand i hand med differentieringsprocessen som ger lätt visualiserade identifierbara steg vilket gör dem att föredra för cellcykelbiologer över, säg modellbakterien E. coli.
beteendet hos däggdjursceller i vävnadsodling har fungerat som grund för mycket av vad vi vet om cellcykeln i högre eukaryoter. Den eukaryota cellcykeln kan i stort sett separeras i två steg, interfas, den del av cellcykeln när cellens material dupliceras och mitos, uppsättningen fysiska processer som deltar i kromosomsegregering och efterföljande celldelning. Hastigheterna för processer i cellcykeln byggs mestadels upp från många av de molekylära händelserna såsom polymerisation av DNA och cytoskeletala filament vars hastigheter vi redan har övervägt. För den karakteristiska cellcykeltiden på 20 timmar i en HeLa-cell ägnas nästan hälften åt G1 (BNID 108483) och nära en annan halv är S-fas (BNID 108485) medan G2 och M är mycket snabbare vid cirka 2-3 timmar respektive 1 timme (BNID 109225, 109226). Det stadium som är mest varierande i varaktighet är G1. I mindre gynnsamma tillväxtförhållanden när cellcykelns varaktighet ökar är detta det stadium som mest påverkas, förmodligen på grund av den tid det tar tills någon reglerande storlekskontroll uppnås. Även om olika typer av bevis pekar på förekomsten av en sådan kontrollpunkt, är det för närvarande mycket dåligt förstått. Historiskt har stadier i cellcykeln vanligtvis härletts med hjälp av fasta celler men nyligen har genetiskt kodade biosensorer som förändrar lokalisering i olika stadier av cellcykeln gjort det möjligt att få levande cell temporal information om cellcykelprogression och arrestering.
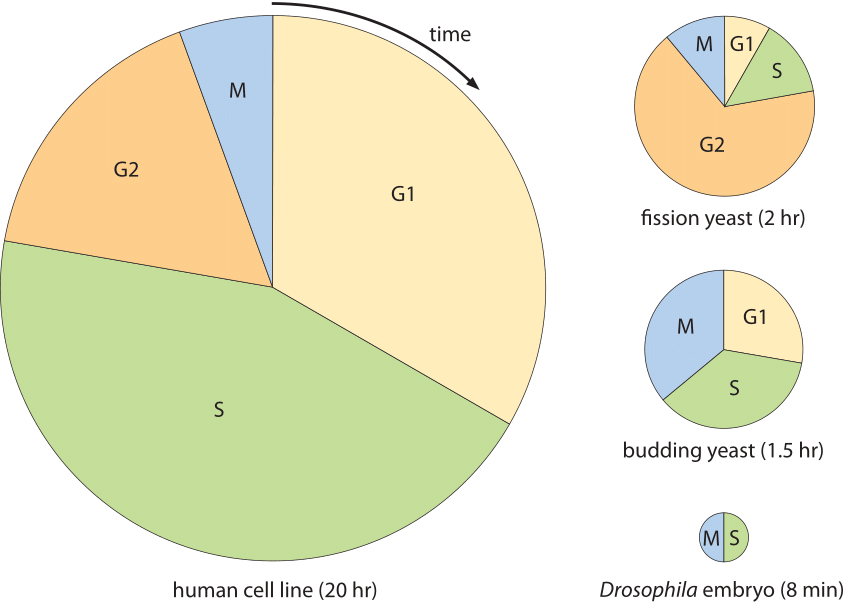
Figur 2: cellcykeltider för olika celltyper. Varje cirkeldiagram visar den del av cellcykeln som ägnas åt var och en av de primära stadierna i cellcykeln. Området för varje diagram är proportionellt mot den totala cellcykelvaraktigheten. Cellcykelvaraktigheter återspeglar minimala fördubblingstider under ideala förhållanden. (Anpassad från” The Cell Cycle – Principles of Control ” av David Morgan.)
hur jämförs cellcykelns längd med den tid det tar en cell att syntetisera sitt nya genom? En frikoppling mellan genomlängden och fördubblingstiden finns i eukaryoter på grund av användningen av flera DNA-replikeringsstartplatser. För däggdjursceller har det observerats att för många vävnader med mycket varierande totala cellcykeltider är varaktigheten av S-fasen där DNA-replikation sker anmärkningsvärt konstant. För musvävnader som de som finns i tjocktarmen eller tungan varierade S-fasen i ett litet intervall från 6, 9 till 7, 5 timmar (BNID 111491). Även vid jämförelse av flera epitelvävnader över människa, råtta, mus och hamster var S-fasen mellan 6 och 8 timmar (BNID 107375). Dessa mätningar utfördes på 1960-talet genom att utföra ett slags pulsjaktförsök med den radioaktivt märkta nukleotidtymidin. Under den korta pulsen införlivades den radioaktiva föreningen endast i genomet av celler i S-fas. Genom att mäta varaktigheten av utseende och sedan försvinnande av märkta celler i m-fas kan man dra slutsatsen hur länge S-fasen varade det faktum att varaktigheten av S-fasen är relativt konstant i sådana celler används till denna dag för att uppskatta cellcykelns varaktighet från en kunskap om endast den fraktion av celler vid en given ögonblicksbild i tid som är i S-fas. Till exempel, om en tredjedel av cellerna ses i S-fas som varar ca 7 timmar, antas cellcykeltiden vara ca 7 timmar/(1/3) 20 timmar. Idag utförs dessa typer av mätningar mestadels med BrdU som markör för S-fas. Vi är inte medvetna om en tillfredsställande förklaring till ursprunget till denna relativt konstanta replikeringstid och hur den är relaterad till hastigheten av DNA-polymeras och densiteten hos replikeringsinitieringsställen längs genomet.
mångfalden av cellcykler visas i Figur 2 och visar flera modellorganismer och varaktigheterna och positioneringen av de olika stadierna i deras cellcykler. Ett extremt exempel förekommer i den fascinerande processen med embryonal utveckling av fruktflugan Drosophila melanogaster. I detta fall skiljer sig situationen från konventionella celldelningar eftersom massan i stället för att syntetisera nya cytoplasmatiska material i huvudsak bevaras förutom replikationen av det genetiska materialet. Detta händer på ett mycket synkront sätt i cirka 10 generationer och en replikationscykel av tusentals celler i embryot, säg mellan cykel 10 och 11, händer på cirka 8 minuter som visas i Figur 2 (BNID 103004, 103005, 110370). Detta är snabbare än replikationstiderna för alla bakterier trots att genomet är 120 miljoner BP lång (BNID 100199). Ett slående exempel på cellernas förmåga att anpassa sin tidsmässiga dynamik.




