Buněčná biologie 07: mikrotubuly a buněčné dělení
toto jsou poznámky z přednášky 7 kurzu buněčné biologie Harvard Extension.
Přednáška 6 představila mikrotubuly a tato přednáška bude diskutovat o jejich roli v buněčném dělení. Zde je úvodní video:
Přehled buněčného cyklu
buněčného cyklu – proces buněčného dělení a replikace se řídí řadu biochemických přepne se nazývá buněčný cyklus řídicího systému.
buněčného cyklu je rozdělen do etap, které jsou rozděleny do fází – lidé se odkazují na „4 fází“, ale pak tam jsou vlastně 5, a lidé také použít jiná slova, aby skupina těch fází dohromady a další slova rozdělit. Snažil jsem se shrnout vztah mezi těmito pojmy v následující tabulce. (upraveno / rozšířeno z Wikipedie):
| MOST general grouping | the supposed „4 phases” | subphases |
|---|---|---|
| non-dividing | Gap 0 (G0) | |
| interphase | Gap 1 (G1) | G1a R G1b |
| Synthesis (S) | ||
| Gap 2 (G2) | ||
| Mitosis | Mitosis (M) | prophase prometaphase metaphase anaphase telophase cytokinesis |
The obsah každé fáze je krásně shrnuta v této vynikající Wikimedia Commons obrázek o Kelvinsong:

nejrychlejší-dělení lidských buněk může dokončit buněčného cyklu v o 24 hodin (G1: 9h, S: 10h, G2: 4h, M: 30 min). Kvasinky mohou dokončit cyklus za 30 minut a nejrychleji se dělící buňky Drosophily trvají jen 8 minut.
Hlavní regulátory tohoto procesu zahrnují cykliny, které regulují cyklin-dependentní kinázu nebo CDK. Připomeňme, že kinázy jsou proteiny, které fosforylují jiné proteiny. Fosforylace cílů CDK umožňuje pokračovat v mitóze. Abych byl přesný, zrání podporu faktor nebo MPF je obligátní heterodimeric komplex složený cyklin B a CDK, který jen dělá svou phosphorylating akce, kdy oba proteiny jsou přítomny.
Role mikrotubulů
mikrotubuly jsou kritické v průběhu buněčného cyklu-organizují buněčné komponenty a rozdělují je na dvě části. Zde je řada videí buněčného cyklu, která zdůrazňují roli mikrotubulů:
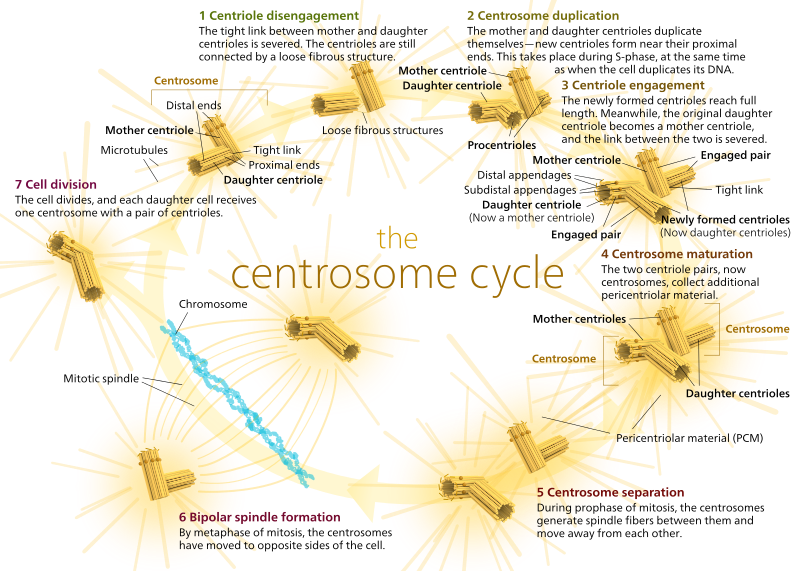
U zvířat, klidový buněk a to i buněk v interfázi mají obvykle jen jeden MTOC, tzv. centrosome, který slouží jako centrální rozbočovač pro všechny mikrotubuly v buňce. A centrosome se skládá ze dvou centriol, jak je uvedeno níže (opět díky Kelvinsong):

dvou centriol odpoutat od sebe a samy replikovat během S fáze, a pak samostatný tvoří opak ‚poláci‘ buňky v M fázi, tak, že teď tam jsou dva MTOCs, z nichž každá bude nakonec jediným MTOC nové buňky (další šéf Kelvinsong obrázek):
Během mitózy, pak máte dvě ‚poláci‘ buňky, každá s mikrotubuly ukotveny na (-) konci a s jejich (+) koncích překrývající, směřující do středu buňky, jak je znázorněno zde (Wikimedia Commons obrázek o Lordjuppiter):
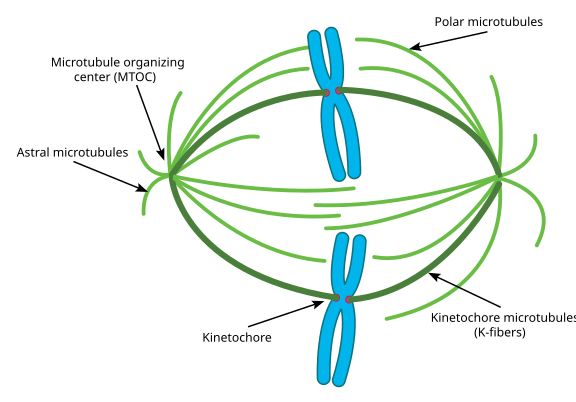
celá věc se nazývá vřetenový aparát a oblast, kde se mikrotubuly obou Mtoc překrývají, se nazývá “ zóna interdigitace.’Někdy uslyšíte každý MTOC a jeho ježkovité pole mikrotubulů zvaných‘ mitotická aster.‘
Mikrotubulů během této fáze jsou řekl, aby spadají do tří kategorií:
- Astrální mikrotubuly bodu směrem ven, směrem do buněk kůry, aby se kotva celé vřeteno přístroje podél osy dělení buněk.
- kinetochorové mikrotubuly se navazují na kinetochor chromatidů.
- polární mikrotubuly, orientované paralelně k sobě, ale v protichůdných směrech, jsou rozhodující pro tlačení vřetenového aparátu během mitózy. (Ve skutečnosti jsou polární mikrotubuly také přítomny dříve a pomáhají tlačit centrosomy od sebe během profáze).
Pokud dáváte přednost fotografií přes diagramy, tady je to, co celé vřeteno přístroje vypadá, se chromatidy v modré, mikrotubuly v zelené, a kinetochores jako červené tečky:

Mikrotubuly se stala mnohem dynamičtější během mitózy: více gama-tubulinu podporuje jednodušší nukleace, ale XMAP215, mikroskopický stabilizátor, je fosforylován a tím inaktivován během mitózy, takže Kinesin-13 zdarma catastrophize mikrotubuly. Bohatství jsou vyrobeny a ztraceny rychle. Poločas mikrotubulů během mitózy je asi 15 minut ve srovnání s 30 minutami během interfáze. Lidé studovat mikrotubulové dynamiky pomocí FRAP: přidat fluorescenční mikroskopický, odbarvit a uvidíte, jak rychle montáží dochází na základě toho, jak brzy fluorescence znovu objeví. + Tipy také hrají hlavní roli při napomáhání a sestavování mikrotubulů.
Kinesin-5 is má dvě polární hlavy, které se váží na protichůdné mikrotubuly a snaží se kráčet směrem k (+) konci každého z nich. To tlačí dva mikrotubuly od sebe a poskytuje hnací sílu pro oddělení Mtoc.
Centromerní DNA má nízkou informační entropii a speciální histony, které se liší od ostatních chromatinů. Centromery jsou jednou částí genomu, kterou téměř nikdy nezachytíte při sekvenování další generace, dokonce i ve opravdu velké hloubce. To proto, že centromery slouží jinému účelu, než hodně ze zbytku genomu: pořadí je příznivé pro interakci s centromeric kinetochorových proteinů a přílohu. Koheziny jsou proteiny, které udržují dvě sesterské chromatidy pohromadě. Budeme odkazovat na kinetochorové proteiny, které mají dvě vrstvy, vnitřní kinetochor a vnější kinetochor.
během prometafázy se chromozomy pohybují tam a zpět. Kinesin kotva chromozomů do kinetochorových mikrotubulů za tip, kde Kinesin-13 je depolymerizing mikrotubuly, podporovaný nedostatek dostupných tubulinových dimerů. Kombinace motorických proteinů, mikrotubulů interagujících proteinů a běhounu slouží k pohybu chromozomů. Mezitím, dynein a dynactin – motorové proteiny, které pěšky směrem k (-) konci – práce na astrální mikrotubuly, tahání MTOCs směrem k periferii buňky. V metafáze se chromatidy vyrovnávají podél „metafázové desky“.
během tohoto procesu se jaderná obálka rozpustí, a tak se jaderný dovoz stává irelevantním. Ran-GEF lokalizuje v blízkosti chromozomů a generuje vysoké koncentrace Ran-GTP, který poskytuje energii pro některé nezbytné procesy (?).
Buňky mají nějaký mechanismus pro detekci napětí v mikrotubulů, která označuje jejich připevnění chromatidy před mitózy může pokračovat. Ujistěte se, že každý chromatid je správně ukotven, je zásadní pro zabránění aneuploidii.
mimochodem, jiné cytoskeletální prvky kromě mikrotubulů také hrají klíčovou roli v buněčném cyklu. Při cytokinezi tvoří aktin kontraktilní kruh a pomocí motorických proteinů myosinu II buňku rozdělí na dvě.
význam modelových organismů
objev regulačních procesů buněčného cyklu se silně spoléhal na některé úhledné rysy populárních modelových organismů.
Saccharomyces cerevisiae (pučící kvasinky) a Schizosaccharomyces pombe (štěpení kvasinky) může existovat jako haploids nebo neznámý. To je důležité, protože v haploidní fázi může jedna mutace vyřadit Gen – nemusíte zasáhnout obě alely. A v droždí, mnoho mutací, zejména v Cdc__ (cell division control) geny, jsou závislé na teplotě, kde protein s missense mutace může stále fungovat správně v „tolerantní“ teploty, ale ztrácí svou nativní funkce na non-tolerantní teploty. To umožňuje studovat vyřazovací fenotyp (při nepovolené teplotě), přičemž má stále pohodlí, že je schopen snadno šířit organismy (při přípustné teplotě). Celý genom s. cerevisiae je k dispozici jako plazmidové knihovny, což umožňuje screening, pro který plazmid zachrání fenotyp daného mutanta. To je, kolik genů, které regulují buněčný cyklus, bylo objeveno.
V S. cerivisiae, nadějné je součástí fáze G1, a jakmile dcera buňka dosáhne určité velikosti, v okamžiku, nazvaný „START“, dva se zavázali vstupem S a nakonec dokončení buněčného cyklu. Savčí buňky mají svůj vlastní závazkový bod nazývaný restrikční bod nebo R, V G1, což je analogické ke spuštění.
mutanti Cdc28 citliví na teplotu se při nepermisivní teplotě nerodí. Že Cdc28 gen kóduje droždí je homolog naše cyklin dependentní kinázy (CDK), které, když a jen když komplexu cyklin, může fosforylují další proteiny, regulovat jejich zapojení do buněčného cyklu fáze. Mutanti citliví na teplotu při nepermisivní teplotě se zaseknou a nemohou vstoupit do fáze S. Místo toho se chovají jako buňky divokého typu zbavené živin: rostou dostatečně velké, aby prošly startem, ale pak nepokračují.
Xenopus (druh žáby) se ukázal jako kritický pro pochopení buněčného cyklu, protože jeho reprodukce zahrnuje velmi velký počet buněk (tj. dostatek výchozího materiálu pro západní bloty atd.), které jsou dokonale synchronizovány (tj. všechny jsou ve stejné fázi buněčného cyklu ve stejném okamžiku. (Ve srovnání s kvasinkami, například, kde buňky nebudou všechny ve stejné fázi ve stejnou dobu). Také samotné vejce je velké a snadno se s ním pracuje a po oplodnění následuje několik buněčných cyklů. V žáby, vejce begin meiotického dělení, ale pak zatčení na G2 fáze po dobu 8 měsíců, zatímco oni rostou ve velikosti a skladovat věci, které budou potřebné pro růst na oplodnění.
Střední filamenta
kromě mikrovlákny a mikrotubulů, eukaryotní buňky mají také řadu „ostatní“ cytoskeletální proteiny zvané střední filamenta (IFs). Ačkoli rozmanitější než mikrofilamenty a mikrotubuly, if nejsou jen termínem pro “ jakékoli jiné vlákno – – spíše jsou skupinou příbuzných proteinů. Obvykle procházejí cytoplazmou a vnitřním jaderným obalem, jsou nepolární a nemají s nimi žádné motorické proteiny. Mají velkou pevnost v tahu a jsou velmi stabilní, s pomalým směnným kurzem a málo členěním, i když fosforylace může podpořit jejich demontáž. Zde jsou některé populární příklady:
- keratiny se nacházejí v epiteliálních buňkách, mezodermálních buňkách a neuronech. Poskytují sílu a přicházejí v kyselých a zásaditých formách. Každý si může vytvořit svůj vlastní pramen, ale většina If se skládá ze dvou pramenů-jednoho zásaditého a jednoho kyselého, jaksi zkrouceného kolem sebe. Vlasy a nehty jsou vyrobeny z „tvrdého“ keratinu bohatého na cystein pro disulfidové vazby, které poskytují nesmírnou sílu. Perms a rovnání se spoléhají na snížení disulfidových vazeb, přetvoření vlasů a následné reformování disulfidových vazeb. Máte také „měkký“ keratin v kůži.
- Desminy, jako je vimentin, se nacházejí v mezenchymálních buňkách (kost, chrupavka a tuk).
- Neurofilamenty jsou v neuronálních axonech a regulují jejich průměr, což zase určuje rychlost šíření akčního potenciálu.
- laminy jsou nejrozšířenější a jsou považovány za nejvíce podobné fylogenetickému předkovi všech ostatních if. Poskytují strukturální podporu pro jadernou membránu. Mohou pomoci vyprostit komplexy jaderných pórů a také uspořádat DNA.
konečně souhrnné video: