Sejtbiológia 07: mikrotubulusok és sejtosztódás
Ezek a Harvard Extension sejtbiológia tanfolyamának 7. előadásának jegyzetei.
a 6.előadás mikrotubulusokat vezetett be, és ez az előadás a sejtosztódásban betöltött szerepüket tárgyalja. Itt van egy bevezető videó:
a sejtciklus áttekintése
a sejtciklust-a sejtosztódás és a replikáció folyamatát-egy sor biokémiai kapcsoló szabályozza, amelyeket sejtciklus-szabályozó rendszernek neveznek.
a sejtciklus fázisokra oszlik, amelyek fázisokra vannak osztva – az emberek a “4 fázisra” utalnak, de valójában 5 van, és az emberek más szavakat is használnak a fázisok csoportosítására, más szavakkal pedig felosztására. Mindent megtettem, hogy a következő táblázatban összefoglaljam a kifejezések közötti kapcsolatot. (módosítva / kibővítve a Wikipédiából):
| MOST general grouping | the supposed “4 phases” | subphases |
|---|---|---|
| non-dividing | Gap 0 (G0) | |
| interphase | Gap 1 (G1) | G1a R G1b |
| Synthesis (S) | ||
| Gap 2 (G2) | ||
| Mitosis | Mitosis (M) | prophase prometaphase metaphase anaphase telophase cytokinesis |
The az egyes fázisok tartalmát gyönyörűen összefoglalja Kelvinsong ezen kiemelkedő Wikimedia Commons képe:

a leggyorsabban osztódó emberi sejtek körülbelül 24 óra alatt képesek befejezni a sejtciklust (G1: 9h, S: 10h, G2: 4h, M: 30 perc). Az élesztő 30 perc alatt képes befejezni a ciklust, és a leggyorsabban osztódó Drosophila sejtek mindössze 8 percet vesznek igénybe.
ennek a folyamatnak a Fővezérlői közé tartoznak a ciklinok, amelyek szabályozzák a ciklinfüggő kinázt vagy a CDK-t. Emlékezzünk arra, hogy a kinázok olyan fehérjék, amelyek más fehérjéket foszforilálnak. A CDK célpontjainak foszforilezése lehetővé teszi a mitózis folytatását. Pontosabban, az érést elősegítő faktor vagy MPF egy kötelező heterodimer komplex, amely ciklin B-ből és CDK-ból áll, amely csak akkor fejti ki foszforiláló hatását, ha mindkét fehérje jelen van.
a mikrotubulusok szerepe
a mikrotubulusok kritikusak a sejtciklus során – sejtkomponenseket szerveznek és kettéosztják őket. Íme egy sor videó a sejtciklusról, amelyek kiemelik a mikrotubulusok szerepét:
állatokban a nyugalmi sejtek, sőt az interfázisban lévő sejtek általában csak egy MTOC-val rendelkeznek, amelyet centroszómának neveznek, amely a sejt összes mikrotubulusának központi csomópontjaként szolgál. A centroszóma két centriolból áll, az alábbiak szerint (ismét a Kelvinsongnak köszönhetően):

a két centriol leválasztódik egymástól, és megismétli magát az S fázisban, majd elkülönül, hogy a sejt ellentétes”pólusait”képezze az M fázisban, így most két Mtoc van, amelyek mindegyike végül egy új sejt egyetlen MTOC-ja lesz (egy másik boss Kelvinsong kép):
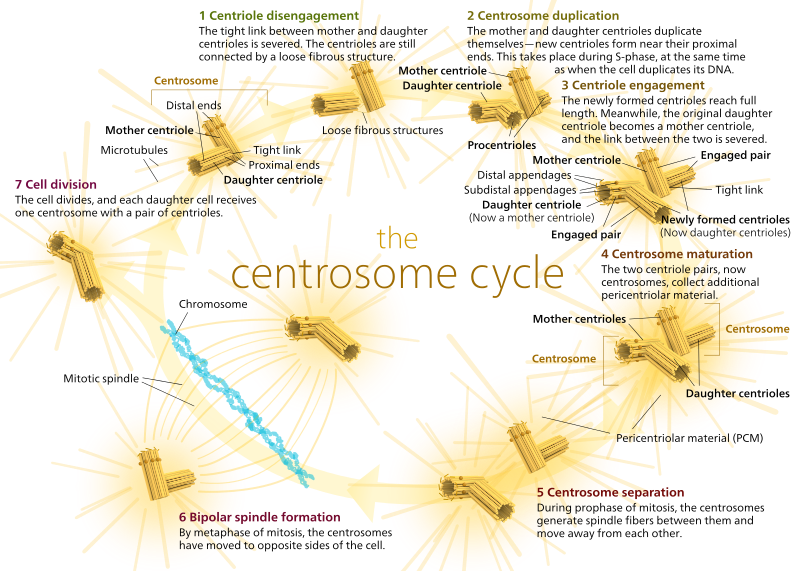
a mitózis során a sejt két “pólusa”van, amelyek mindegyike mikrotubulusokkal van rögzítve a (-) végén, és a (+) végeik átfedésben vannak, a cella közepére mutatnak, amint az itt látható (Wikimedia Commons kép: Lordjuppiter):
ezt az egészet orsókészüléknek nevezzük, és azt a területet, ahol a két Mtoc mikrotubulus átfedésben van, interdigitációs zónának nevezzük. Néha hallani fogsz minden egyes MTOC-t és annak sünszerű mikrotubulus-tömbjét, amit mitotikus aster-nek hívnak.’
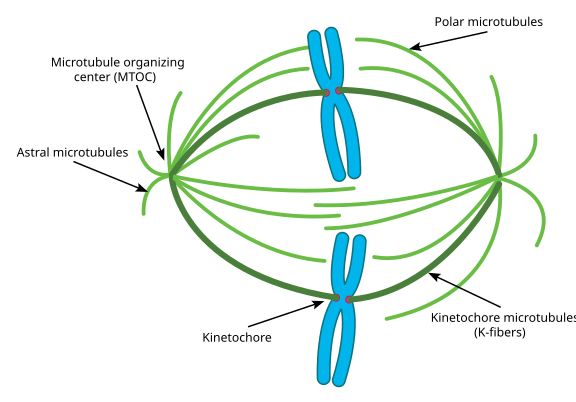
a mikrotubulusok ebben a szakaszban állítólag három kategóriába sorolhatók:
- az asztrális mikrotubulusok kifelé mutatnak, a sejtkéreg felé, annak érdekében, hogy az egész orsókészüléket a sejtosztódás tengelye mentén rögzítsék.
- Kinetochore mikrotubulusok kapcsolódnak a kromatidok kinetochorához.
- az egymással párhuzamosan, de ellentétes irányban orientált poláris mikrotubulusok döntő fontosságúak az orsókészüléknek a mitózis során történő szétnyomásához. (Valójában a poláris mikrotubulusok korábban is jelen vannak, és segítik a centroszómákat a profázis során).
Ha jobban szereted a fényképeket a diagramokkal szemben, így néz ki az egész orsókészülék, kék kromatidokkal, zöld mikrotubulusokkal, a kinetochores pedig piros pontokkal:

a mikrotubulusok sokkal dinamikusabbá válnak a mitózis során: több gamma-tubulin elősegíti a könnyebb nukleációt, de az XMAP215, egy mikrotubulus stabilizátor, foszforilálódik és így inaktiválódik a mitózis során, így a kinezin-13 szabadon hagyja a mikrotubulusokat. A vagyon gyorsan el is fogy. A mikrotubulus felezési ideje a mitózis során körülbelül 15 perc, szemben az interfázis alatti 30 perccel. Az emberek a Frap segítségével tanulmányozzák a mikrotubulus dinamikáját: fluoreszkáló mikrotubulust adnak hozzá, fehérítik, és megnézik, hogy milyen gyorsan történik az összeszerelés annak alapján, hogy a fluoreszcencia milyen hamar jelenik meg újra. + A tippek szintén fontos szerepet játszanak a mikrotubulusok összeállításában.
a kinezin-5-nek két poláris feje van, amelyek az ellentétes mikrotubulusokhoz kötődnek, és megpróbálnak mindegyik (+) vége felé járni. Ez tolja a két mikrotubulusok egymástól, és biztosítja a hajtóerőt a szétválasztása a Mtoc.
a centromer DNS alacsony információs entrópiával és speciális hisztonokkal rendelkezik, amelyek különböznek a többi kromatintól. A centromerek a genom egyik része, amelyet szinte soha nem vesz fel a következő generációs szekvenálás során, még igazán nagy mélységben is. Ez azért van, mert a centromerek más célt szolgálnak, mint a genom többi része: az ottani szekvencia kedvező a centromer fehérjékkel való kölcsönhatáshoz és a kinetochore kötődéshez. A kohézinek olyan fehérjék, amelyek együtt tartják a két testvérkromatidot. A kinetochore fehérjékre úgy hivatkozunk, hogy két rétegük van, a belső kinetochore és a külső kinetochore.
a prometafázis során a kromoszómák előre-hátra mozognak. A kinezinek a kromoszómákat a kinetochore mikrotubulusokhoz rögzítik a csúcson túl, ahol a kinezin-13 depolimerizálja a mikrotubulusokat, a rendelkezésre álló tubulin dimerek hiányával segítve. A motoros fehérjék, a mikrotubulus kölcsönhatásban lévő fehérjék és a futópad kombinációja a kromoszómák mozgatását szolgálja. Eközben a dynein és a dynactin – motoros fehérjék, amelyek a (-) vég felé haladnak, az asztrális mikrotubulusokon dolgoznak, az Mtoc-kat a sejt perifériája felé húzva. A metafázisban a kromatidok a metafázis lemez mentén igazodnak.
e folyamat során a nukleáris burok feloldódik, így a nukleáris import irrelevánssá válik. A ran-GEF a kromoszómák közelében helyezkedik el, és nagy koncentrációban termel Ran-GTP-t, amely energiát biztosít néhány szükséges folyamathoz (?).
a sejteknek van valamilyen mechanizmusuk a mikrotubulusok feszültségének kimutatására, amely jelzi kötődési kromatidjaikat, mielőtt a mitózis folytatódhat. Az aneuploidia elkerüléséhez elengedhetetlen annak biztosítása, hogy minden kromatid megfelelően rögzítve legyen.
egyébként a mikrotubulusokon kívül más citoszkeletális elemek is kulcsszerepet játszanak a sejtciklusban. A citokinezisben az aktin kontraktilis gyűrűt képez, és a miozin II motoros fehérjék segítségével a sejtet kettévágja.
a modellszervezetek fontossága
a sejtciklus szabályozási folyamatainak felfedezése nagymértékben támaszkodott a népszerű modellszervezetek néhány ügyes tulajdonságára.
a Saccharomyces cerevisiae (kezdő élesztő) és a Schizosaccharomyces pombe (hasadási élesztő) létezhet haploidként vagy diploidként. Ez azért fontos, mert a haploid fázisban egy mutáció kiüthet egy gént – nem kell mindkét allélt eltalálnia. Az élesztőben sok mutáció, különösen a Cdc__ (cell division control) génekben, hőmérsékletfüggő, ahol a missense mutációval rendelkező fehérje továbbra is megfelelően működhet ‘megengedő’ hőmérsékleten, de elveszíti natív funkcióját ‘nem megengedő’ hőmérsékleten. Ez lehetővé teszi a kiütéses fenotípus tanulmányozását (nem megengedő hőmérsékleten), miközben továbbra is kényelmesen képes az organizmusok könnyű szaporítására (megengedő hőmérsékleten). A teljes S. cerevisiae Genom plazmidkönyvtárakként áll rendelkezésre, lehetővé téve annak szűrését, hogy melyik plazmid megmenti az adott mutáns fenotípusát. Ez az, hogy hány olyan gént fedeztek fel, amelyek szabályozzák a sejtciklust.
S-Ben. cerivisiae, a bimbózás a G1 fázis része, és amint a leánysejt elér egy bizonyos méretet, egy “START” – nak nevezett pillanatban, a kettő elkötelezi magát az S-be való belépés és végül a sejtciklus befejezése mellett. Az emlős sejteknek megvan a saját elkötelezettségi pontjuk, amelyet restrikciós pontnak vagy R-nek hívnak, a G1-ben, amely analóg a START-val.
hőmérséklet-érzékeny Cdc28 mutánsok nem bimbóznak a nem megengedő hőmérsékleten. A Cdc28 gén kódolja a ciklinfüggő kináz (CDK) élesztő homológját, amely akkor és csak akkor képes foszforilálni más fehérjéket, hogy szabályozza részvételüket a sejtciklus fázisaiban. A hőmérséklet-érzékeny mutánsok a nem megengedő hőmérsékleten elakadnak, nem tudnak rügyezni és belépni az S fázisba. Ehelyett úgy viselkednek, mint a tápanyagoktól megfosztott vad típusú sejtek: elég nagyok ahhoz, hogy átmenjenek a Starton, de aztán nem folytatják.
a Xenopus (egyfajta béka) kritikusnak bizonyult a sejtciklus megértése szempontjából, mivel szaporodása nagyon sok sejtet (pl. elég kiindulási anyag a Western blotokhoz stb.), amelyek tökéletesen szinkronizálva vannak (azaz mindegyik ugyanabban a fázisban van sejtciklus ugyanabban a pillanatban. (Hasonlítsuk össze például az élesztővel, ahol a sejtek nem lesznek ugyanabban a fázisban ugyanabban az időben). Maga a tojás is nagy és könnyen kezelhető, és több sejtciklus követi a megtermékenyítést. A békákban a tojások megkezdik a meiotikus osztódást, majd a G2 fázisban 8 hónapig megállnak, miközben méretük növekszik, és felhalmozódnak a megtermékenyítés után a növekedéshez szükséges dolgok.
közbenső szálak
a mikrofilamentumokon és mikrotubulusokon kívül az eukarióta sejtek számos más citoszkeletális fehérjével is rendelkeznek, amelyeket közbenső szálaknak (IFs) neveznek. Bár változatosabbak, mint a mikrofilamentumok és a mikrotubulusok, Az IFs nem csak egy catch-all kifejezés a ‘bármely más izzószálra’, hanem a kapcsolódó fehérjék egy csoportja. Általában a citoplazmán és a belső magburokon keresztül terjednek, nem polárisak, és nem kapcsolódnak hozzájuk motoros fehérjék. Nagy szakítószilárdsággal rendelkeznek, és nagyon stabilak, lassú árfolyammal és nem sok bontással, bár a foszforiláció elősegítheti szétszerelésüket. Íme néhány népszerű példa:
- a keratinok hámsejtekben, mezodermasejtekben és neuronokban találhatók. Erősséget biztosítanak, savas és bázikus formában jönnek létre. Mindegyik saját szálat alkothat, de a legtöbb IFs két szálból áll – egy bázikus és egy savas, egyfajta csavart egymás körül. A haj és a köröm kemény keratinból készül, amely ciszteinben gazdag diszulfidkötésekhez, ami biztosítja a hatalmas erőt. A Perms és az egyenesítés a diszulfidkötések csökkentésére, a haj átalakítására, majd a diszulfidkötések megreformálására támaszkodik. A bőrödben is van’ puha ‘ keratin.
- a Dezminok, mint például a vimentin, megtalálhatók a mesenchymalis sejtekben (csont, cartiledge és zsír).
- a Neurofilamentumok neuronális axonokban vannak, és szabályozzák azok átmérőjét, ami viszont meghatározza az akciós potenciál terjedésének sebességét.a
- Laminok mind a legelterjedtebbek, és úgy gondolják, hogy leginkább hasonlítanak az összes többi IFs filogenetikai őséhez. Szerkezeti támogatást nyújtanak a nukleáris membrán számára. Segíthetnek a nukleáris póruskomplexumok elhelyezésében és a DNS szervezésében is.
végül egy összefoglaló videó: