Concetti di biologia-1a edizione canadese
I virus furono scoperti per la prima volta dopo lo sviluppo di un filtro in porcellana, chiamato filtro Chamberland-Pasteur, che poteva rimuovere tutti i batteri visibili al microscopio da qualsiasi campione liquido. Nel 1886, Adolph Meyer dimostrò che una malattia delle piante di tabacco, la malattia del mosaico del tabacco, poteva essere trasferita da una pianta malata a una sana attraverso estratti vegetali liquidi. Nel 1892, Dmitri Ivanowski dimostrò che questa malattia poteva essere trasmessa in questo modo anche dopo che il filtro Chamberland-Pasteur aveva rimosso tutti i batteri vitali dall’estratto. Tuttavia, sono passati molti anni prima che fosse dimostrato che questi agenti infettivi “filtrabili” non erano semplicemente batteri molto piccoli ma erano un nuovo tipo di minuscole particelle che causano malattie.
I virioni, singole particelle virali, sono molto piccoli, circa 20-250 nanometri (1 nanometro = 1/1.000. 000 mm). Queste singole particelle virali sono la forma infettiva di un virus al di fuori della cellula ospite. A differenza dei batteri (che sono circa 100 volte più grandi), non possiamo vedere i virus con un microscopio ottico, ad eccezione di alcuni grandi virioni della famiglia dei poxvirus (Figura 12.3).
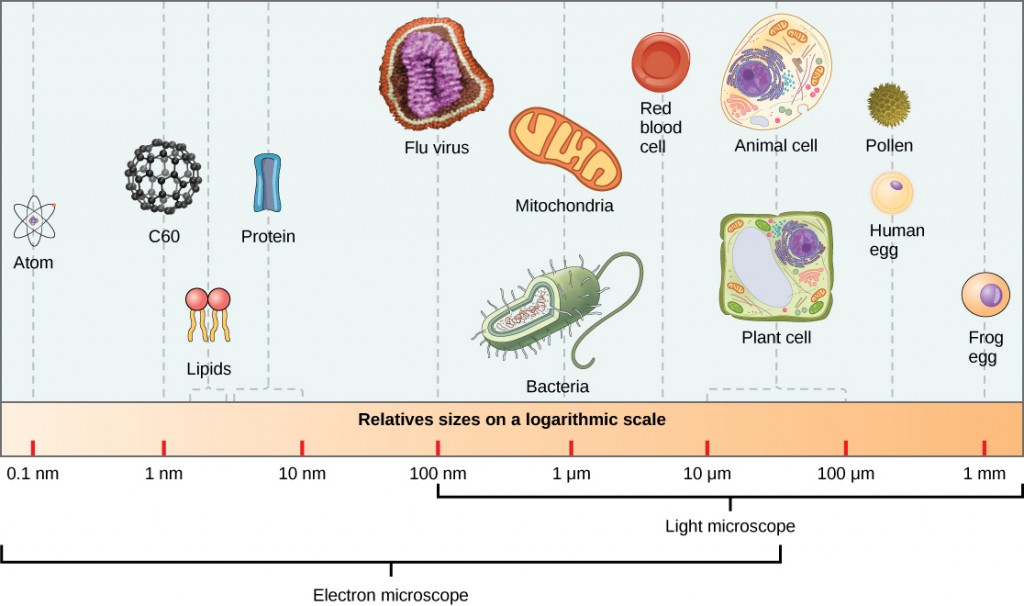
Non fu fino allo sviluppo del microscopio elettronico negli anni ‘ 40 che gli scienziati ottennero la loro prima buona visione della struttura del virus del mosaico del tabacco (Figura 12.2) e altri. La struttura superficiale dei virioni può essere osservata sia mediante microscopia elettronica a scansione che a trasmissione, mentre le strutture interne del virus possono essere osservate solo in immagini da un microscopio elettronico a trasmissione (Figura 12.4).
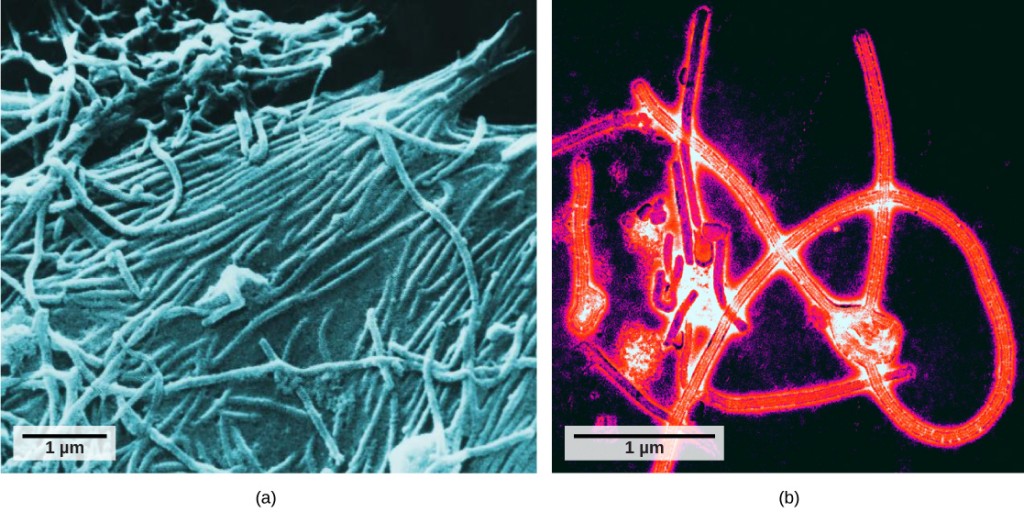
L’utilizzo di questa tecnologia ha permesso la scoperta di molti virus di tutti i tipi di organismi viventi. Sono stati inizialmente raggruppati per morfologia condivisa, il che significa la loro dimensione, forma e strutture distintive. Successivamente, i gruppi di virus sono stati classificati in base al tipo di acido nucleico che contenevano, DNA o RNA e se il loro acido nucleico era a singolo o doppio filamento. Più recentemente, l’analisi molecolare dei cicli di replicazione virale ha ulteriormente affinato la loro classificazione.
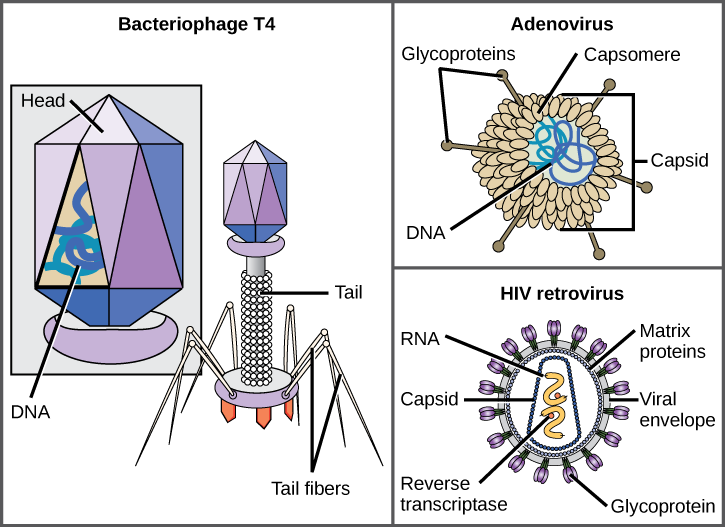
Un virione è costituito da un nucleo di acido nucleico, un rivestimento proteico esterno e talvolta un involucro esterno costituito da membrane proteiche e fosfolipidi derivate dalla cellula ospite. La differenza più visibile tra i membri delle famiglie virali è la loro morfologia, che è piuttosto diversa. Una caratteristica interessante della complessità virale è che la complessità dell’ospite non è correlata alla complessità del virione. Alcune delle strutture virioniche più complesse sono osservate nei batteriofagi, virus che infettano gli organismi viventi più semplici, i batteri.
I virus sono disponibili in molte forme e dimensioni, ma questi sono coerenti e distinti per ogni famiglia virale (Figura 12.5). Tutti i virioni hanno un genoma di acido nucleico coperto da uno strato protettivo di proteine, chiamato capside. Il capside è costituito da subunità proteiche chiamate capsomeri. Alcuni capsidi virali sono semplici “sfere” poliedriche, mentre altri sono piuttosto complessi nella struttura. La struttura esterna che circonda il capside di alcuni virus è chiamata busta virale. Tutti i virus usano una sorta di glicoproteina per attaccarsi alle loro cellule ospiti a molecole sulla cellula chiamate recettori virali. Il virus sfrutta queste molecole di superficie cellulare, che la cellula utilizza per qualche altro scopo, come un modo per riconoscere e infettare specifici tipi di cellule. Ad esempio, il virus del morbillo utilizza una glicoproteina di superficie cellulare nell’uomo che normalmente funziona nelle reazioni immunitarie e possibilmente nell’interazione sperma-uovo alla fecondazione. L’attaccamento è un requisito per i virus per penetrare successivamente nella membrana cellulare, iniettare il genoma virale e completare la loro replicazione all’interno della cellula.
Il batteriofago T4, che infetta il batterio E. coli, è tra i virioni più complessi conosciuti; T4 ha una struttura di coda proteica che il virus utilizza per attaccarsi alla cellula ospite e una struttura della testa che ospita il suo DNA.
L’adenovirus, un virus animale non evoluto che causa malattie respiratorie negli esseri umani, utilizza picchi proteici che sporgono dai suoi capsomeri per attaccarsi alla cellula ospite. I virus non inveloped includono anche quelli che causano la poliomielite (poliovirus), le verruche plantari (papillomavirus) e l’epatite A (virus dell’epatite A). I virus non Inveloped tendono ad essere più robusti e più probabilità di sopravvivere in condizioni difficili, come l’intestino.
I virioni avvolti come l’HIV (virus dell’immunodeficienza umana), l’agente eziologico dell’AIDS (sindrome da immunodeficienza acquisita), sono costituiti da acido nucleico (RNA nel caso dell’HIV) e proteine del capside circondate da un involucro a doppio strato di fosfolipidi e dalle sue proteine associate (Figura 12.5). Varicella, influenza e parotite sono esempi di malattie causate da virus con buste. A causa della fragilità dell’involucro, i virus non inviluppati sono più resistenti ai cambiamenti di temperatura, pH e alcuni disinfettanti rispetto ai virus avviluppati.
Nel complesso, la forma del virione e la presenza o l’assenza di un involucro ci dice poco su quali malattie i virus possono causare o quali specie potrebbero infettare, ma è ancora un mezzo utile per iniziare la classificazione virale.
Quale delle seguenti affermazioni sulla struttura del virus è vera?
A) Tutti i virus sono racchiusi in una membrana virale.
B) Il capsomero è costituito da piccole subunità proteiche chiamate capsidi.
C) Il DNA è il materiale genetico di tutti i virus.
D) Le glicoproteine aiutano il virus ad attaccarsi alla cellula ospite.
<!- D – >
A differenza di tutti gli organismi viventi che utilizzano il DNA come materiale genetico, i virus possono utilizzare DNA o RNA come loro. Il nucleo del virus contiene il genoma o il contenuto genetico totale del virus. I genomi virali tendono ad essere piccoli rispetto ai batteri o agli eucarioti, contenenti solo quei geni che codificano per le proteine che il virus non può ottenere dalla cellula ospite. Questo materiale genetico può essere a filamento singolo o a doppio filamento. Può anche essere lineare o circolare. Mentre la maggior parte dei virus contiene un singolo segmento di acido nucleico, altri hanno genomi composti da diversi segmenti.
I virus a DNA hanno un nucleo di DNA. Il DNA virale dirige le proteine della replica della cellula ospite per sintetizzare le nuove copie del genoma virale e per trascrivere e tradurre quel genoma nelle proteine virali. I virus del DNA causano malattie umane come la varicella, l’epatite B e alcune malattie veneree come l’herpes e le verruche genitali.
I virus a RNA contengono solo RNA nei loro nuclei. Per replicare i loro genomi nella cellula ospite, i genomi dei virus RNA codificano enzimi non presenti nelle cellule ospiti. Gli enzimi della RNA polimerasi non sono stabili come le DNA polimerasi e spesso commettono errori durante la trascrizione. Per questo motivo, mutazioni, cambiamenti nella sequenza nucleotidica, nei virus a RNA si verificano più frequentemente che nei virus a DNA. Ciò porta a un’evoluzione e un cambiamento più rapidi nei virus a RNA. Ad esempio, il fatto che l’influenza sia un virus a RNA è uno dei motivi per cui ogni anno è necessario un nuovo vaccino antinfluenzale. Le malattie umane causate da virus a RNA includono l’epatite C, il morbillo e la rabbia.
I virus possono essere visti come parassiti intracellulari obbligati. Il virus deve attaccarsi a una cellula vivente, essere preso all’interno, fabbricare le sue proteine e copiare il suo genoma, e trovare un modo per sfuggire alla cellula in modo che il virus possa infettare altre cellule e, infine, altri individui. I virus possono infettare solo alcune specie di ospiti e solo alcune cellule all’interno di quell’ospite. La base molecolare per questa specificità è che una particolare molecola di superficie, nota come recettore virale, deve essere trovata sulla superficie della cellula ospite affinché il virus si attacchi. Inoltre, le differenze metaboliche osservate in diversi tipi di cellule basate sull’espressione genica differenziale sono un fattore probabile in cui le cellule che un virus può utilizzare per replicarsi. La cellula deve fare le sostanze di cui il virus ha bisogno, come gli enzimi per cui il genoma del virus stesso non ha geni, o il virus non sarà in grado di replicarsi usando quella cellula.
Fasi delle infezioni da virus
Un virus deve “prendere in consegna” una cellula per replicarsi. Il ciclo di replicazione virale può produrre cambiamenti biochimici e strutturali drammatici nella cellula ospite, che possono causare danni alle cellule. Questi cambiamenti, chiamati effetti citopatici, possono cambiare le funzioni cellulari o addirittura distruggere la cellula. Alcune cellule infette, come quelle infettate dal virus del raffreddore comune (rhinovirus), muoiono per lisi (scoppio) o apoptosi (morte cellulare programmata o “suicidio cellulare”), rilasciando tutti i virioni della progenie contemporaneamente. I sintomi delle malattie virali derivano dalla risposta immunitaria al virus, che tenta di controllare ed eliminare il virus dal corpo e dal danno cellulare causato dal virus. Molti virus animali, come l’HIV (virus dell’immunodeficienza umana), lasciano le cellule infette del sistema immunitario mediante un processo noto come germogliamento, in cui i virioni lasciano la cellula individualmente. Durante il processo di germogliamento, la cellula non subisce la lisi e non viene immediatamente uccisa. Tuttavia, il danno alle cellule che l’HIV infetta può rendere impossibile per le cellule di funzionare come mediatori di immunità, anche se le cellule rimangono in vita per un periodo di tempo. Le infezioni virali più produttive seguono passaggi simili nel ciclo di replicazione del virus: allegato, penetrazione, svolgimento, replica, assemblaggio e rilascio.
Un virus si attacca a uno specifico sito recettoriale sulla membrana cellulare ospite attraverso proteine di attacco nel capside o proteine incorporate nel suo involucro. L’attaccamento è specifico, e in genere un virus si attacca solo alle cellule di una o poche specie e solo alcuni tipi di cellule all’interno di quelle specie con i recettori appropriati.
Concetto in azione

Guarda questo video per una spiegazione visiva di come l’HIV e l’influenza attaccano il corpo.
A differenza dei virus animali, l’acido nucleico dei batteriofagi viene iniettato nella cellula ospite nuda, lasciando il capside all’esterno della cellula. I virus vegetali e animali possono entrare nelle loro cellule attraverso l’endocitosi, in cui la membrana cellulare circonda e avvolge l’intero virus. Alcuni virus avvolti entrano nella cellula quando l’involucro virale si fonde direttamente con la membrana cellulare. Una volta all’interno della cellula, il capside virale viene degradato e l’acido nucleico virale viene rilasciato, che diventa quindi disponibile per la replicazione e la trascrizione.
Il meccanismo di replicazione dipende dal genoma virale. I virus del DNA di solito usano proteine ed enzimi delle cellule ospiti per produrre DNA aggiuntivo che viene utilizzato per copiare il genoma o essere trascritto in RNA messaggero (mRNA), che viene quindi utilizzato nella sintesi proteica. I virus a RNA, come il virus dell’influenza, di solito usano il nucleo di RNA come modello per la sintesi di RNA genomico virale e mRNA. L’mRNA virale viene tradotto in enzimi virali e proteine del capside per assemblare nuovi virioni (Figura 12.6). Naturalmente, ci sono eccezioni a questo modello. Se una cellula ospite non fornisce gli enzimi necessari per la replicazione virale, i geni virali forniscono le informazioni per la sintesi diretta delle proteine mancanti. I retrovirus, come l’HIV, hanno un genoma a RNA che deve essere trascritto al contrario per produrre DNA, che poi viene inserito nel DNA dell’ospite. Per convertire l’RNA in DNA, i retrovirus contengono geni che codificano la trascrittasi inversa dell’enzima specifico del virus che trascrive un modello di RNA in DNA. Il fatto che l’HIV produca alcuni dei propri enzimi, che non si trovano nell’ospite, ha permesso ai ricercatori di sviluppare farmaci che inibiscono questi enzimi. Questi farmaci, incluso l’inibitore della trascrittasi inversa AZT, inibiscono la replicazione dell’HIV riducendo l’attività dell’enzima senza influenzare il metabolismo dell’ospite.
L’ultimo stadio della replicazione virale è il rilascio dei nuovi virioni nell’organismo ospite, dove sono in grado di infettare le cellule adiacenti e ripetere il ciclo di replicazione. Alcuni virus vengono rilasciati quando la cellula ospite muore e altri virus possono lasciare le cellule infette germogliando attraverso la membrana senza uccidere direttamente la cellula.
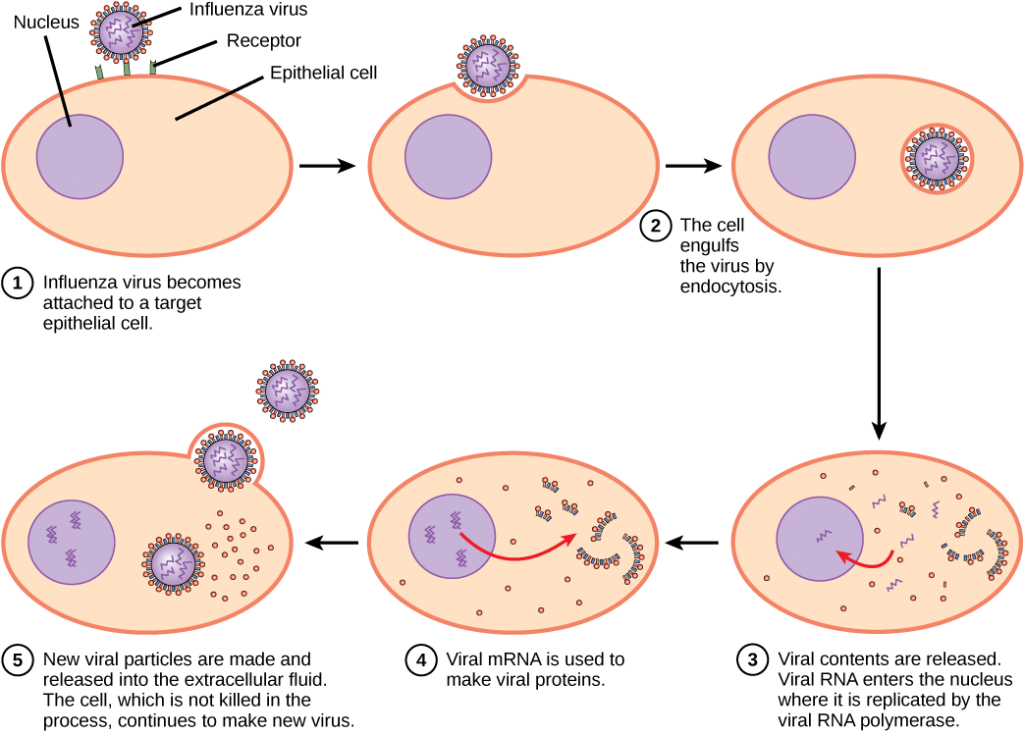
Il virus dell’influenza è confezionato in un involucro virale, che si fonde con la membrana plasmatica. In questo modo, il virus può uscire dalla cellula ospite senza ucciderlo. Quale vantaggio ottiene il virus mantenendo viva la cellula ospite?
<!- La cellula ospite può continuare a produrre nuove particelle virali.- >
Concetto in azione

Fai clic su questo tutorial sui virus per identificare strutture, modalità di trasmissione, replica e altro ancora.