細胞生物学07:微小管と細胞分裂
これらは、ハーバード大学エクステンションの細胞生物学コースの講義7からのノートです。
第6講義では微小管を紹介し、この講義では細胞分裂におけるそれらの役割について議論します。 ここでは紹介ビデオです:
細胞周期の概要
細胞周期-細胞分裂と複製のプロセス–は、細胞周期制御システムと呼ばれる一連の生化学的スイッチに
細胞周期は、フェーズに分割されたフェーズに分割されています–人々は”4つのフェーズ”を参照しますが、実際には5があり、人々はまた、それらのフェーズを一緒にグループ化するために他の言葉を使用し、それらを細分化するために他の言葉を使用します。 これらの用語間の関係を次の表に要約するために最善を尽くしました。 (Wikipediaから変更/拡張):
| MOST general grouping | the supposed “4 phases” | subphases |
|---|---|---|
| non-dividing | Gap 0 (G0) | |
| interphase | Gap 1 (G1) | G1a R G1b |
| Synthesis (S) | ||
| Gap 2 (G2) | ||
| Mitosis | Mitosis (M) | prophase prometaphase metaphase anaphase telophase cytokinesis |
The 各段階の内容は、Kelvinsongによるこの優れたウィキメディアコモンズの画像に美しく要約されています:

最速の分裂ヒト細胞は、約24時間(G1:9h、S:10h、G2:4h、M:30分)で細胞周期を完了することができる。 酵母は30分でサイクルを終了することができ、最速分裂ショウジョウバエ細胞はわずか8分かかります。
このプロセスのマスターコントローラには、サイクリン依存性キナーゼまたはCDKを調節するサイクリンが含まれる。 キナーゼは他のタンパク質をリン酸化するタンパク質であることを思い出してください。 その標的のCDKのリン酸化は、有糸分裂が進行することを可能にする。 正確には、成熟促進因子またはMPFは、サイクリンBとCDKからなる偏性ヘテロ二量体複合体であり、両方のタンパク質が存在する場合にのみリン酸化作用
微小管の役割
微小管は、細胞周期全体にわたって重要であり、細胞成分を組織し、それらを二つに分割する。 ここでは、微小管の役割を強調する細胞周期のビデオのシリーズがあります:動物では、静止細胞や間期の細胞でさえ、通常、細胞内のすべての微小管の中心ハブとして機能する中心体と呼ばれるMTOCを1つだけ持っています。
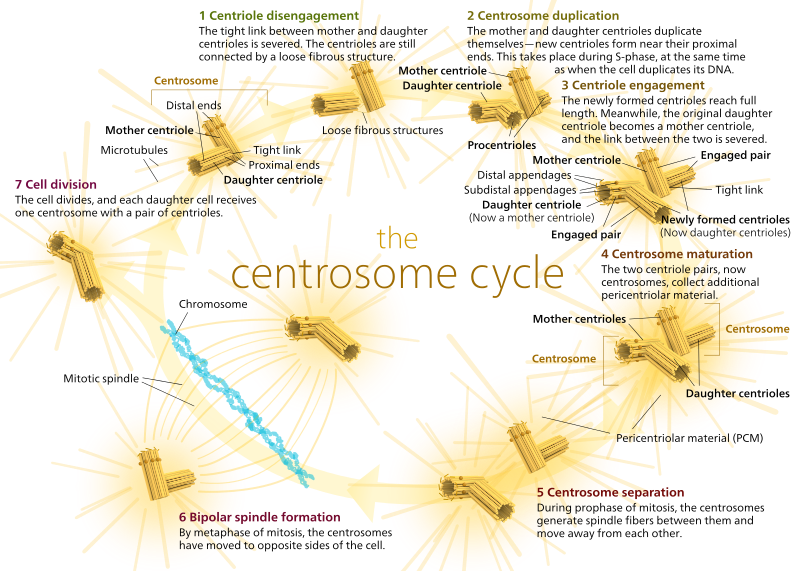
中心体は、以下に示すように二つの中心小体で構成されています(Kelvinsongに再び感謝します):

二つの中心小体は互いに離脱し、S期の間に自分自身を複製し、M期の間に細胞の反対の”極”を形成するために分離し、二つのMtocがあり、それぞれが最終的に新しい細胞(別のボスKelvinsong画像)の唯一のMTOCとなる。
有糸分裂の間、細胞の二つの”極”があり、それぞれが微小管が(-)端に固定され、それらの(+)端が重なって、ここに示すように細胞の中心を指しています(Wikimedia Commons image by Lordjuppiter):
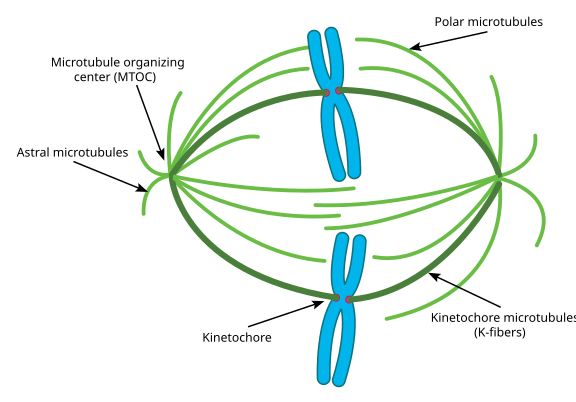
その全部をスピンドル装置と呼び、二つのMtocの微小管が重なる領域を”interdigitationのゾーン”と呼びます。”あなたは時々、各MTOCと”有糸分裂aster”と呼ばれる微小管のそのウニのような配列を聞くでしょう。’
この段階の微小管は、三つのカテゴリに分類されると言われています:
- アストラル微小管は、細胞分裂の軸に沿ってスピンドル装置全体を固定するために、細胞皮質に向かって外側を指しています。
- 動原体微小管は、染色分体の動原体に付着する。
- 極性微小管は、互いに平行であるが対向する方向に配向し、有糸分裂中にスピンドル装置を離れて押すために重要である。 (実際には、極性微小管も早期に存在し、前期の間に中心体を離れてプッシュするのに役立ちます)。
図よりも写真を好む場合は、スピンドル装置全体がどのように見えるかを示します。

微小管は有糸分裂の間にはるかに動的になります:より多くのガンマチューブリンはより容易な核形成を促進しますが、微小管安定剤であるXMAP215は有糸分裂の間にリン酸化され、したがって不活性化され、キネシン-13は微小管を破壊するのに自由になります。 運命は作られ、すぐに失われます。 有糸分裂の間の微小管の半減期は、間期の間の30分と比較して約15分である。 人々はFRAPを使用して微小管のダイナミクスを研究します:蛍光微小管を追加し、それを漂白し、蛍光がどのくらい早く再び現れるかに基づいて再組 +ヒントはまた、微小管を支援し、組み立てる上で主要な役割を果たしています。キネシン-5は、対向する微小管に結合し、それぞれの(+)端に向かって歩こうとする二つの極性頭部を有する。 これは、二つの微小管を離れてプッシュし、Mtocの分離のための駆動力を提供します。
動原体DNAは、情報エントロピーが低く、他のクロマチンとは異なる特殊なヒストンを有する。 動原体は、本当に高い深さであっても、次世代シークエンシングではほとんど拾うことのないゲノムの一部です。 動原体は、ゲノムの残りの部分の多くとは異なる目的を提供しているためです:そこの配列は、動原体タンパク質と動原体付着との相互作用のために好 コヒーシンは、2つの姉妹染色分体を一緒に保つタンパク質です。 我々は、二つの層、内側の動原体と外側の動原体を有するものとして動原体タンパク質を参照します。
前後期の間、染色体は前後に移動します。 キネシンは、キネシン-13が利用可能なチューブリン二量体の不足によって助け、微小管を解重合されている先端を超えてキネトコア微小管に染色体を モーター蛋白質、微小管相互作用蛋白質およびtreadmillingの組合せは染色体を動かすのに役立ちます。 一方、(–)エンドに向かって歩くダイニンとダイナクチン-モータータンパク質は、細胞周辺に向かってMtocを引っ張って、アストラル微小管上で動作します。 中期では、染色分体は”中期プレート”に沿って整列するようになる。
このプロセスの間、核エンベロープは溶解するので、核の輸入は無関係になります。 Ran-GEFは染色体の近くに局在化し、いくつかの必要なプロセスのためのエネルギーを提供するRan-GTPの高濃度を生成します(?).
細胞は、有糸分裂が進行する前に、それらの付着染色分体を示す微小管の張力を検出するためのいくつかのメカニズムを有する。
細胞は、有糸分裂を進行させることができる。 すべての染色分体が適切に固定されていることを確認することは、異数性を回避するために重要です。
ところで、微小管以外の他の細胞骨格要素も細胞周期において重要な役割を果たす。 細胞質分裂では、アクチンは収縮リングを形成し、ミオシンIIモータータンパク質の助けを借りて細胞を二つに収縮させる。
モデル生物の重要性
細胞周期調節プロセスの発見は、一般的なモデル生物のいくつかのきちんとした特徴に大きく依存していました。Saccharomyces cerevisiae(出芽酵母)およびSchizosaccharomyces pombe(分裂酵母)は、一倍体または二倍体として存在することができます。
一倍体期では、1つの突然変異が遺伝子をノックアウトする可能性があるため、両方の対立遺伝子をヒットする必要はないため、これは重要です。 酵母では、多くの突然変異、特にCdc__(細胞分裂制御)遺伝子では、ミスセンス変異を有するタンパク質は依然として”許容”温度で適切に機能するが、”非許容”温度でその本来の機能を失う温度依存性である。 これにより、(許容温度で)生物を容易に増殖させることができるという利便性を有しながら、ノックアウト表現型を(非許容温度で)研究することが可 S.cerevisiaeゲノム全体がプラスミドライブラリーとして利用可能であり、どのプラスミドが与えられた変異体の表現型を救助するかをスクリーニングすることが可能である。 それは、細胞周期を調節する遺伝子のいくつが発見されたかです。Sで
cerivisiae、出芽は相G1の一部であり、娘細胞が一定のサイズに達すると、”開始”と呼ばれる瞬間に、二人はSに入り、最終的に細胞周期を完了することにコミット 哺乳動物細胞は、G1において、制限点またはRと呼ばれる独自のコミットメント点を有し、これは開始に類似する。
温度感受性Cdc28変異体は、非透過的な温度で発芽しません。
温度感受性Cdc28変異体は、非透過的な温度で発芽しません。 Cdc28遺伝子は、サイクリン依存性キナーゼ(CDK)の酵母のホモログをコードしており、サイクリンと複合体化した場合にのみ、他のタンパク質をリン酸化して細胞周期期への関与を調節することができる。 非透過温度での温度感受性変異体は、芽を出してS期に入ることができない立ち往生します。 代わりに、彼らは栄養素を奪われた野生型細胞のように振る舞う:彼らは開始を通過するのに十分な大きさに成長するが、その後継続しない。
アフリカツメガエル(カエルの一種)は、その再生が非常に多数の細胞を含むため、細胞周期を理解するために重要であることが証明された(すなわち、 西部のしみ、等のための十分な出発材料。 完全に同期されている(すなわち、すべてが同じ瞬間に細胞周期の同じ段階にある)。 (例えば、細胞がすべて同時に同じ段階にあるわけではない酵母と比較して)。 また、卵自体は大きく、作業が容易であり、複数の細胞周期が受精に続く。 カエルでは、卵は減数分裂を開始しますが、g2期で8ヶ月間停止し、受精時に成長に必要なものを貯蔵します。
中間フィラメント
マイクロフィラメントと微小管に加えて、真核細胞はまた、中間フィラメント(IFs)と呼ばれる”他の”細胞骨格タンパク質のホス マイクロフィラメントや微小管よりも多様ですが、IFsは単に”他のフィラメント”のキャッチオール用語ではなく、関連するタンパク質のグループです。 それらは一般に、細胞質および内部核エンベロープを通って延び、非極性であり、それらに関連するモータータンパク質を有さない。 リン酸化が分解を促進できるけれどもそれらに大きい引張強さがあり、遅い為替レートおよび多くの故障と非常に安定しています。 ここではいくつかの一般的な例があります:
- ケラチンは、上皮細胞、中胚葉細胞およびニューロンに見出される。 それらは強さを提供し、酸性および基本的な形態入って来。 それぞれが独自の鎖を形成することができますが、ほとんどのIfは二つの鎖で構成されています–一つの塩基性と一つの酸性、お互いの周りにねじれ 髪と爪は、巨大な強さを提供するジスルフィド結合のためのシステインが豊富な”ハード”ケラチンで作られています。 パーマおよびまっすぐになることはジスルフィドの結束を減らし、毛を造り直し、次にジスルフィドの結束を改良することに頼ります。 また、あなたの肌に”柔らかい”ケラチンがあります。
- ビメンチンなどのデスミンは、間葉系細胞(骨、軟骨および脂肪)に見出される。
- ニューロフィラメントは神経軸索にあり、その直径を調節し、活動電位伝播の速度を決定する。
- ラミンは両方とも最も広く普及しており、他のすべてのIfの系統発生の祖先に最も類似していると考えられています。 それらは核膜の構造的支持を提供する。 彼らは核孔複合体をスペースアウトし、またDNAを整理するのに役立つかもしれません。
最後に、要約ビデオ: