Concepts of Biology–1st Canadian Edition
ウイルスは、Chamberland-Pasteurフィルターと呼ばれる磁器フィルターの開発後に最初に発見されました。 1886年、アドルフ-マイヤーは、タバコ植物の病気、タバコモザイク病は、液体植物抽出物を介して病気の植物から健康な植物に移すことができることを実証した。 1892年、Dmitri Ivanowskiは、Chamberland-Pasteurフィルターが抽出物からすべての生菌を除去した後でも、この病気がこのように伝染する可能性があることを示しました。 それでも、これらの「濾過可能な」感染性病原体は単に非常に小さな細菌ではなく、小さな病気を引き起こす新しいタイプの粒子であることが証明されたのは何年も前のことでした。
単一のウイルス粒子であるビリオンは非常に小さく、約20-250ナノメートル(1ナノメートル=1/1,000,000mm)である。 これらの個々のウイルス粒子は、宿主細胞外のウイルスの感染性形態である。 細菌(約100倍の大きさ)とは異なり、ポックスウィルスファミリーのいくつかの大きなビリオンを除いて、光学顕微鏡でウイルスを見ることはできません(図12.3)。
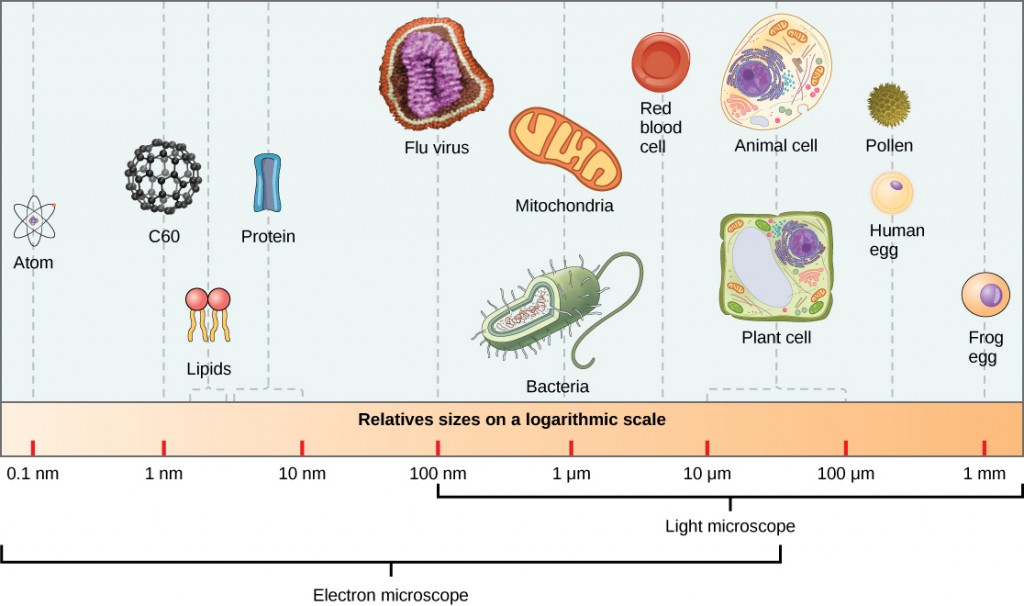
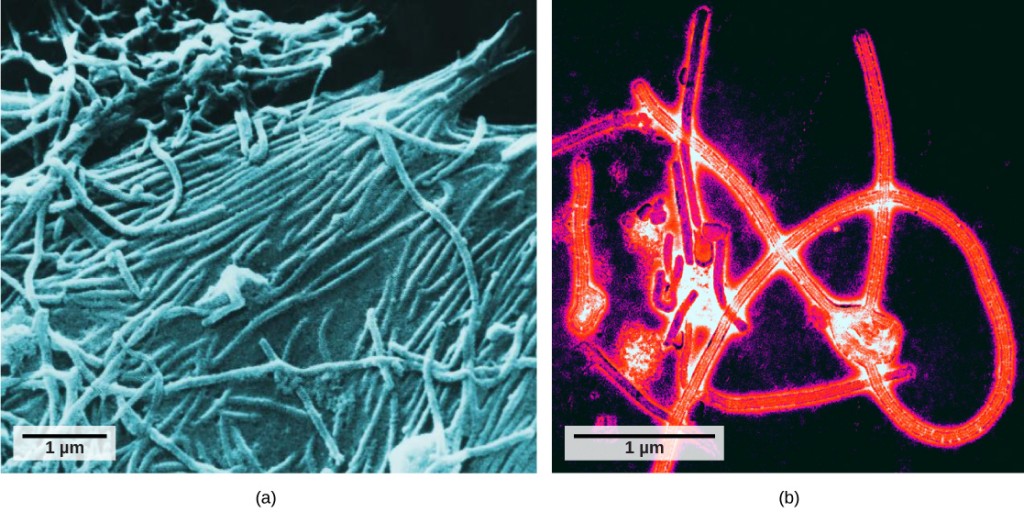
科学者がタバコモザイクウイルス(図12.2)などの構造を最初によく見たのは、1940年代の電子顕微鏡の開発までではありませんでした。 ビリオンの表面構造は走査型電子顕微鏡と透過型電子顕微鏡の両方で観察することができますが、ウイルスの内部構造は透過型電子顕微鏡からの画像でのみ観察することができます(図12.4)。
この技術の使用は、すべてのタイプの生物の多くのウイルスの発見を可能にした。 彼らは当初、その大きさ、形状、および区別構造を意味し、共有形態によってグループ化されました。 その後、ウイルスの群は、それらが含有する核酸の種類、DNAまたはRNA、およびそれらの核酸が一本鎖または二本鎖であるかどうかによって分類された。 より最近では、ウイルス複製サイクルの分子分析は、さらにその分類を洗練しています。
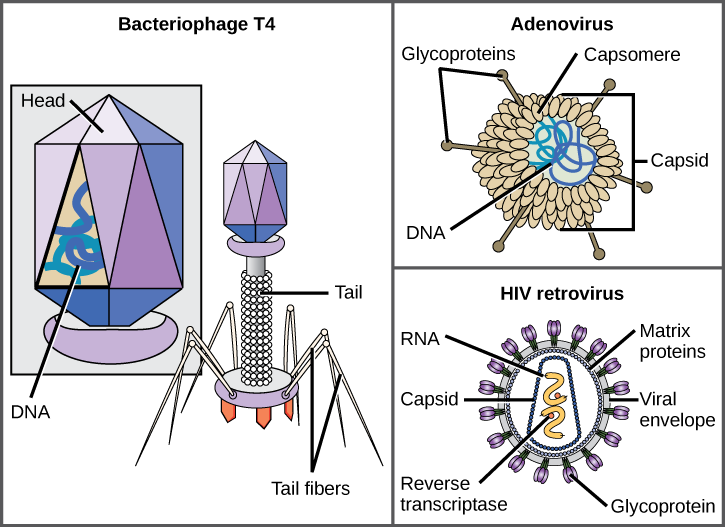
ビリオンは、核酸コア、外側のタンパク質コーティング、および宿主細胞に由来するタンパク質およびリン脂質膜からなる外側のエンベロープからなる。 ウイルス家族のメンバー間の最も目に見える違いは、その形態であり、これは非常に多様である。 ウイルスの複雑さの興味深い特徴は、宿主の複雑さがビリオンの複雑さと相関しないことである。 最も複雑なビリオン構造のいくつかは、バクテリオファージ、最も単純な生物、細菌に感染するウイルスで観察される。
ウイルスには多くの形や大きさがありますが、これらはウイルスファミリーごとに一貫しており、異なるものです(図12.5)。 すべてのビリオンは、カプシドと呼ばれるタンパク質の保護層によって覆われた核酸ゲノムを有する。 カプシドは、カプソメアと呼ばれるタンパク質サブユニットで作られています。 他の人が構造が非常に複雑であるのに対し、いくつかのウイルスのカプシドは、単純な多面体の”球”です。 いくつかのウイルスのカプシドを囲む外側の構造は、ウイルスエンベロープと呼ばれています。 すべてのウイルスはウイルスの受容器と呼出される細胞の分子で宿主細胞に付すのに一種の糖蛋白質を使用します。 このウイルスは、特定の細胞型を認識して感染させる方法として、細胞が他の目的のために使用するこれらの細胞表面分子を利用する。 例えば、麻疹ウイルスは、通常、免疫反応およびおそらく受精時の精子-卵の相互作用において機能するヒトの細胞表面糖タンパク質を使用する。 付着は、ウイルスが後に細胞膜に浸透し、ウイルスゲノムを注入し、細胞内でそれらの複製を完了するための要件である。
大腸菌に感染するT4バクテリオファージは、知られている最も複雑なビリオンの一つであり、T4は、ウイルスが宿主細胞に付着するために使用す
アデノウイルス、ヒトの呼吸器疾患を引き起こす非enveloped動物ウイルスは、宿主細胞に付着するために、そのカプソメアから突出したタンパク質スパイクを使用しています。 非envelopedウイルスには、ポリオ(ポリオウイルス)、足底疣贅(パピローマウイルス)、およびa型肝炎(A型肝炎ウイルス)を引き起こすウイルスも含まれます。 Nonenvelopedウイルスは腸のような粗い条件の下で、存続してがより強く、本当らしい傾向があります。
エイズ(後天性免疫不全症候群)の原因物質であるHIV(ヒト免疫不全ウイルス)のようなエンベロープされたビリオンは、核酸(HIVの場合はRNA)とリン脂質二層エンベロープとそれに関連するタンパク質によって囲まれたカプシドタンパク質からなる(図12.5)。 水痘、インフルエンザおよび流行性耳下腺炎は封筒が付いているウイルスによって引き起こされる病気の例です。 エンベロープの脆弱性のために、非エンベロープウイルスは、エンベロープウイルスよりも温度、pH、およびいくつかの消毒剤の変化に対してより耐性があ
全体的に、ビリオンの形状とエンベロープの有無は、ウイルスが引き起こす可能性のある病気や感染する可能性のある種についてはほとんど教えてくれませんが、ウイルス分類を開始するための有用な手段です。
ウイルス構造に関する以下のステートメントのどれが真ですか?
A)すべてのウイルスはウイルス膜に包まれています。
B)capsomereはcapsidsと呼ばれる小さなタンパク質サブユニットで構成されています。
C)DNAはすべてのウイルスの遺伝物質です。
D)糖タンパク質は、ウイルスが宿主細胞に付着するのを助ける。
D)糖タンパク質は、ウイルスが宿主細胞に付着するのp>
<!–D->
DNAを遺伝物質として使用するすべての生物とは異なり、ウイルスはDNAまたはRNAのいずれかを彼らのものとして使用することが ウイルスコアには、ウイルスのゲノムまたは総遺伝的内容が含まれています。 ウイルスゲノムは細菌や真核生物と比較して小さくなる傾向があり、ウイルスが宿主細胞から得ることができないタンパク質をコードする遺伝子 この遺伝物質は、一本鎖または二本鎖であり得る。 それはまた、線形または円形であってもよい。 ほとんどのウイルスは核酸の単一のセグメントを含んでいますが、他のウイルスはいくつかのセグメントからなるゲノムを持っています。DNAウイルスはDNAコアを持っています。
ウイルスDNAはウイルスのゲノムの新しいコピーを総合し、ウイルス蛋白質にそのゲノムを転写し、翻訳するために宿主細胞の複製蛋白質を指示します。 DNAウイルスは、水痘、b型肝炎、およびヘルペスや生殖器疣贅のようないくつかの性病などの人間の病気を引き起こします。RNAウイルスはRNAのみをコアに含んでいます。
宿主細胞内でそれらのゲノムを複製するために、RNAウイルスのゲノムは宿主細胞には見られない酵素をコードする。 RNAポリメラーゼ酵素はDNAポリメラーゼほど安定ではなく、転写中にミスをすることが多い。 このため、RNAウイルスにおける変異、塩基配列の変化は、DNAウイルスよりも頻繁に起こる。 これは、RNAウイルスのより急速な進化と変化につながります。 例えば、インフルエンザがRNAウイルスであるという事実は、毎年新しいインフルエンザワクチンが必要とされる理由の1つです。 RNAウイルスによって引き起こされる人間の病気には、C型肝炎、麻疹、狂犬病が含まれます。
ウイルスは偏性細胞内寄生虫として見ることができます。 ウイルスは、生きた細胞に付着し、内部に取り込まれ、そのタンパク質を製造し、そのゲノムをコピーし、ウイルスが他の細胞および最終的に他の個体に感染することができるように、細胞を脱出する方法を見つけなければなりません。 ウイルスは、特定の種の宿主とその宿主内の特定の細胞のみに感染する可能性があります。 この特異性の分子基盤は、ウイルス受容体として知られる特定の表面分子が、ウイルスが付着するために宿主細胞表面上に見出されなければならな また、差動遺伝子発現に基づく異なる細胞型で見られる代謝の違いは、ウイルスが複製するために細胞を使用する可能性のある因子である。 細胞は、ウイルスゲノム自体が遺伝子を持たない酵素など、ウイルスが必要とする物質を作らなければならない、またはウイルスはその細胞を使用して複製することができない。
ウイルス感染の手順
ウイルスは複製するために細胞を”引き継ぐ”必要があります。 ウイルスの複製周期は細胞の損傷を引き起こすかもしれない宿主細胞の劇的な生化学的な、構造変更を作り出すことができます。 細胞変性効果と呼ばれるこれらの変化は、細胞機能を変化させたり、細胞を破壊したりする可能性があります。 風邪ウイルス(ライノウイルス)に感染した細胞のようないくつかの感染細胞は、溶解(破裂)またはアポトーシス(プログラムされた細胞死または”細胞自殺”) ウイルス性疾患の症状は、ウイルスに対する免疫応答に起因し、ウイルスを体内から制御および排除しようとするウイルス、およびウイルスによっ HIV(ヒト免疫不全ウイルス)のような多くの動物ウイルスは、出芽として知られるプロセスによって免疫系の感染細胞を去り、そこでビリオンは細胞を個別に離れる。 出芽プロセスの間、細胞は溶解を受けず、すぐには死滅しない。 しかし、HIVに感染する細胞への損傷は、細胞が一定期間生きているにもかかわらず、細胞が免疫のメディエーターとして機能することを不可能にするかも ほとんどの生産性の高いウイルス感染は、ウイルス複製サイクルで同様の手順に従います: 添付ファイル、浸透、コーティング解除、複製、アセンブリ、およびリリース。
ウイルスは、カプシド内の付着タンパク質またはそのエンベロープに埋め込まれたタンパク質を介して宿主細胞膜上の特定の受容体部位に付着 付着は特異的であり、典型的には、ウイルスは、1種または数種の細胞にのみ付着し、適切な受容体を有するそれらの種内の特定の細胞型のみに付着す
アクションの概念

HIVとインフルエンザが体を攻撃する方法の視覚的な説明動物ウイルスとは異なり、バクテリオファージの核酸は裸で宿主細胞に注入され、カプシドは細胞の外に残される。
動物ウイルスとは異なり、バクテリオファージの核酸は宿主細胞の外に注入される。 植物および動物のウイルスは細胞膜が全体のウイルスを囲み、巻き込むendocytosisによって細胞に入ることができます。 ウイルスエンベロープが細胞膜と直接融合すると、いくつかのエンベロープウイルスが細胞に入る。 細胞内に入ると、ウイルスカプシドが分解され、ウイルス核酸が放出され、複製および転写のために利用可能になる。
複製機構はウイルスゲノムに依存する。 DNAウイルスは、通常、宿主細胞タンパク質および酵素を使用して、ゲノムをコピーするために使用される追加のDNAを作成するか、またはメッセンジャー RNA(mRNA)に転写され、タンパク質合成に使用されます。 インフルエンザウイルスのようなRNAウイルスは、通常、ウイルスゲノムRNAおよびmRNAの合成のための鋳型としてRNAコアを使用する。 ウイルスmRNAは、ウイルス酵素およびカプシドタンパク質に翻訳され、新しいビリオンを組み立てる(図12.6)。 もちろん、このパターンには例外があります。 宿主細胞がウイルス複製に必要な酵素を提供しない場合、ウイルス遺伝子は不足しているタンパク質の直接合成に情報を提供する。 HIVのようなレトロウイルスは、dnaを作るために逆転写されなければならないRNAゲノムを有し、それは宿主のDNAに挿入される。 RNAをDNAに変換するために、レトロウイルスは、RNA鋳型をDNAに転写するウイルス特異的酵素逆転写酵素をコードする遺伝子を含む。 HIVが宿主には見られない独自の酵素のいくつかを産生するという事実は、研究者がこれらの酵素を阻害する薬物を開発することを可能にした。 逆転写酵素阻害剤AZTを含むこれらの薬物は、宿主の代謝に影響を与えることなく酵素の活性を低下させることによってHIV複製を阻害する。
ウイルス複製の最後の段階は、新しいビリオンが宿主生物に放出され、隣接する細胞に感染し、複製サイクルを繰り返すことができることである。 いくつかのウイルスは、宿主細胞が死ぬと放出され、他のウイルスは、細胞を直接殺すことなく膜を通って出芽することによって感染細胞を残すこ
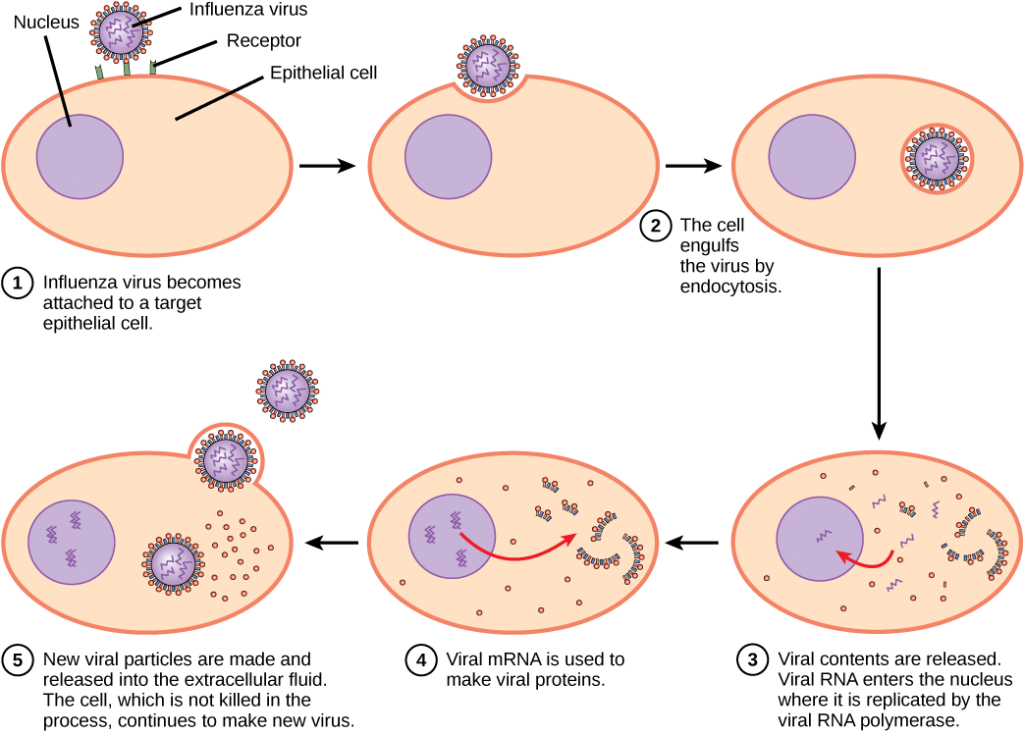
インフルエンザウイルスは、原形質膜と融合するウイルスエンベロープにパッケージ化されています。 このようにして、ウイルスは宿主細胞を殺すことなく宿主細胞を終了することができる。 宿主細胞を生きたままにすることによって、ウイルスはどのような利点を得るのですか?p>
<>
アクションの概念

構造、伝送、複製、およびより多くのモードを識別するために、ウイ
div