Biología Celular 07: Microtúbulos y División Celular
Estas son notas de la conferencia 7 del curso de Biología Celular de Harvard Extension.
La conferencia 6 introdujo los microtúbulos, y esta conferencia discutirá su papel en la división celular. Aquí hay un video introductorio:
Descripción general del ciclo celular
El ciclo celular, el proceso de división y replicación celular, se rige por una serie de interruptores bioquímicos llamados sistema de control del ciclo celular.
El ciclo celular se divide en fases que se dividen en fases: la gente se referirá a las «4 fases», pero en realidad hay 5, y la gente también usa otras palabras para agrupar esas fases y otras palabras para subdividirlas. He hecho todo lo posible para resumir la relación entre estos términos en la siguiente tabla. (modificado / expandido de Wikipedia):
| MOST general grouping | the supposed «4 phases” | subphases |
|---|---|---|
| non-dividing | Gap 0 (G0) | |
| interphase | Gap 1 (G1) | G1a R G1b |
| Synthesis (S) | ||
| Gap 2 (G2) | ||
| Mitosis | Mitosis (M) | prophase prometaphase metaphase anaphase telophase cytokinesis |
The el contenido de cada fase está bellamente resumido en esta excelente imagen de Wikimedia Commons de Kelvinsong:

Las células humanas de división más rápida pueden completar un ciclo celular en aproximadamente 24 horas (G1: 9h, S: 10h, G2: 4h, M: 30 min). La levadura puede terminar un ciclo en 30 minutos, y las células de Drosophila que se dividen más rápido tardan tan solo 8 minutos.
Los controladores maestros de este proceso incluyen las ciclinas, que regulan la quinasa dependiente de ciclina o CDK. Recuerde que las quinasas son proteínas que fosforilan otras proteínas. La fosforilación de CDK de sus blancos permite que la mitosis proceda. Para ser precisos, el factor promotor de maduración o MPF es un complejo heterodimérico obligado compuesto por ciclina B y CDK, que solo realiza su acción fosforiladora cuando ambas proteínas están presentes.
Papel de los microtúbulos
Los microtúbulos son críticos a lo largo del ciclo celular: organizan los componentes celulares y los dividen en dos. Aquí hay una serie de videos del ciclo celular que destacan el papel de los microtúbulos:
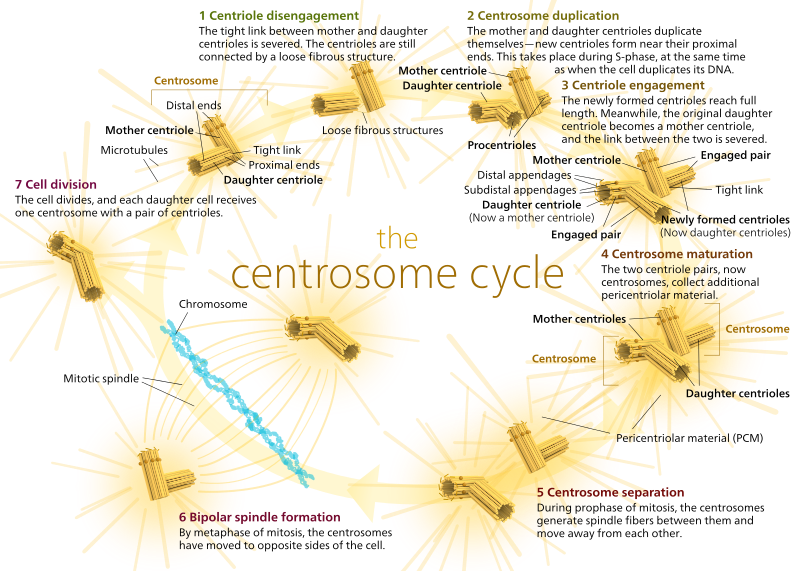
En los animales, las células quiescentes e incluso las células en interfase generalmente tienen solo un COT, llamado centrosoma, que sirve como el eje central para todos los microtúbulos de la célula. Un centrosoma se compone de dos centriolos como se muestra a continuación (gracias de nuevo a Kelvinsong):

Los dos centríolos se desenganchan entre sí y se replican durante la fase S, y luego se separan para formar polos opuestos de la célula durante la fase M, de modo que ahora hay dos MTOCs, cada uno de los cuales eventualmente será el único MTOC de una nueva célula (otra imagen de boss Kelvinsong):
Durante la mitosis, entonces, tienes los dos ‘polos’ de la célula, cada uno con microtúbulos anclados en el extremo (-) y con sus extremos (+) superpuestos, apuntando al centro de la célula, como se muestra aquí (imagen de Wikimedia Commons de Lordjuppiter):
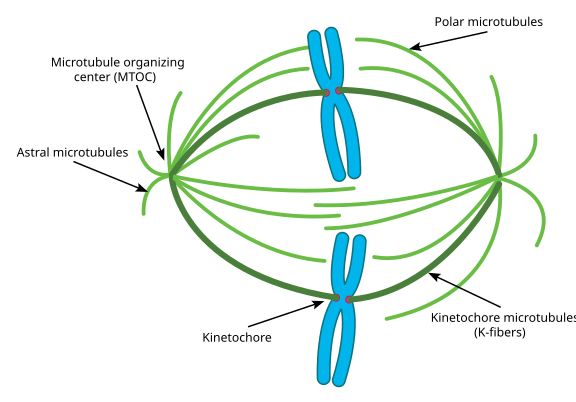
Todo esto se llama un aparato de huso, y el área donde los microtúbulos de los dos MTOC se superponen se llama «zona de interdigitación». A veces oirás cada COT y su conjunto de microtúbulos en forma de erizo llamado aster mitótico.Los microtúbulos ‘
durante esta etapa se dice que caen en tres categorías:
- Los microtúbulos astrales apuntan hacia afuera, hacia la corteza celular, para anclar todo el aparato de huso a lo largo del eje de división celular.
- Los microtúbulos del cinetocoro se unen al cinetocoro de las cromátidas.
- Los microtúbulos polares, orientados paralelos entre sí pero en direcciones opuestas, son cruciales para separar el aparato del huso durante la mitosis. (De hecho, los microtúbulos polares también están presentes antes y ayudan a separar los centrosomas durante la profase).
Si prefiere fotos en lugar de diagramas, así es como se ve todo el aparato de husillo, con cromátidas en azul, microtúbulos en verde y los cinetocoros como puntos rojos:

Los microtúbulos se vuelven mucho más dinámicos durante la mitosis: más gamma-tubulina promueve una nucleación más fácil, pero XMAP215, un estabilizador de microtúbulos, se fosforila y, por lo tanto, se inactiva durante la mitosis, dejando libre a la quinesina-13 para catastrofizar los microtúbulos. Las fortunas se hacen y se pierden rápidamente. La vida media de un microtúbulo durante la mitosis es de aproximadamente 15 minutos, en comparación con 30 minutos durante la interfase. Las personas estudian la dinámica de los microtúbulos usando FRAP: agregue un microtúbulo fluorescente, blanquéelo y vea qué tan rápido se está volviendo a ensamblar en función de qué tan pronto reaparece la fluorescencia. + Las puntas también juegan un papel importante en ayudar y ensamblar los microtúbulos.
Kinesin-5 tiene dos cabezas polares que se unen a microtúbulos opuestos y tratan de caminar hacia el extremo (+) de cada uno. Esto separa los dos microtúbulos y proporciona la fuerza motriz para la separación de los MTOC.
El ADN centromérico tiene baja entropía de información e histonas especiales que difieren de otras cromatinas. Los centrómeros son una parte del genoma que casi nunca se recoge en la secuenciación de próxima generación, incluso a una profundidad muy alta. Esto se debe a que los centrómeros tienen un propósito diferente que gran parte del resto del genoma: la secuencia allí es favorable para la interacción con las proteínas centroméricas y la unión de cinetocoros. Las cohesinas son proteínas que mantienen unidas las dos cromátidas hermanas. Nos referiremos a las proteínas del cinetocoro como que tienen dos capas, el cinetocoro interno y el cinetocoro externo.
Durante la prometafase, los cromosomas se mueven hacia adelante y hacia atrás. Las cinesinas anclan los cromosomas a los microtúbulos del cinetocoro más allá de la punta donde la cinesina-13 despolimeriza los microtúbulos, ayudados por la escasez de dímeros de tubulina disponibles. Una combinación de proteínas motoras, proteínas que interactúan con los microtúbulos y cintas de correr sirve para mover los cromosomas. Mientras tanto, las proteínas motoras de dineína y dinactina que caminan hacia el extremo ( – ) trabajan en los microtúbulos astrales, tirando de los MTOC hacia la periferia celular. En metafase, las cromátidas se alinean a lo largo de la «placa de metafase».
Durante este proceso, la envoltura nuclear se disuelve y así importación nuclear se vuelve irrelevante. Ran-GEF localiza cromosomas cercanos y genera altas concentraciones de Ran-GTP que proporciona energía para algunos procesos necesarios (?).Las células
tienen algún mecanismo para detectar la tensión en los microtúbulos que indica sus cromátidas de fijación antes de que la mitosis pueda proceder. Asegurarse de que cada cromátida esté correctamente anclada es crucial para evitar la aneuploidía.
Por cierto, otros elementos citoesqueléticos además de los microtúbulos también juegan un papel clave en el ciclo celular. En la citocinesis, la actina forma un anillo contráctil y, con la ayuda de proteínas motoras de miosina II, cincha la célula en dos.
Importancia de los organismos modelo
El descubrimiento de los procesos reguladores del ciclo celular se basó en gran medida en algunas características nítidas de los organismos modelo populares.
Saccharomyces cerevisiae (levadura en ciernes) y Schizosaccharomyces pombe (levadura de fisión) pueden existir como haploides o diploides. Esto es importante porque en la fase haploide, una mutación puede eliminar un gen; no es necesario golpear ambos alelos. Y en la levadura, muchas mutaciones, especialmente en los genes Cdc__ (control de división celular), dependen de la temperatura, donde una proteína con una mutación sin sentido todavía puede funcionar correctamente a temperaturas «permisivas», pero pierde su función nativa a temperaturas «no permisivas». Esto hace posible estudiar el fenotipo knockout (a la temperatura no permisiva) sin dejar de tener la conveniencia de poder propagar fácilmente los organismos (a la temperatura permisiva). Todo el genoma de S. cerevisiae está disponible como bibliotecas de plásmidos, lo que permite seleccionar para qué plásmido rescata el fenotipo de un mutante dado. Así es como se descubrieron muchos de los genes que regulan el ciclo celular.
En S. cerivisiae, el brote es parte de la fase G1, y una vez que la célula hija alcanza un cierto tamaño, en un momento llamado «INICIO», las dos se comprometen a entrar en S y, finalmente, completar el ciclo celular. Las células de mamíferos tienen su propio punto de compromiso llamado punto de restricción o R, en G1, que es análogo a START.
Los mutantes Cdc28 sensibles a la temperatura no brotan a la temperatura no permisiva. El gen Cdc28 codifica el homólogo de levadura de nuestra quinasa dependiente de ciclina (CDK) que, cuando y solo cuando se complica con ciclina, puede fosforilar otras proteínas para regular su participación en las fases del ciclo celular. Los mutantes sensibles a la temperatura a la temperatura no permisiva se atascan incapaces de brotar y entrar en la fase S. En cambio, se comportan como células de tipo salvaje sin nutrientes: crecen lo suficientemente grandes como para comenzar pero luego no continúan.
Xenopus (una especie de rana) demostró ser fundamental para comprender el ciclo celular, ya que su reproducción involucra un gran número de células (p.ej. suficiente material de partida para manchas occidentales, etc.) que están perfectamente sincronizados (es decir, todos están en la misma fase del ciclo celular en el mismo momento. (Compare con la levadura, por ejemplo, donde las células no estarán todas en la misma fase al mismo tiempo). También el óvulo en sí es grande y fácil de trabajar, y múltiples ciclos celulares siguen la fertilización. En las ranas, los huevos comienzan la división meiótica, pero luego se detienen en la fase G2 durante 8 meses mientras crecen en tamaño y acumulan las cosas que se necesitarán para el crecimiento después de la fertilización.
Filamentos intermedios
Además de microfilamentos y microtúbulos, las células eucariotas también tienen una gran cantidad de «otras» proteínas citoesqueléticas llamadas filamentos intermedios (IFs). Aunque son más diversos que los microfilamentos y microtúbulos, los IFs no son solo un término general para «cualquier otro filamento», sino que son un grupo de proteínas relacionadas. Generalmente se extienden a través del citoplasma y la envoltura nuclear interna, no son polares y no tienen proteínas motoras asociadas con ellos. Tienen una gran resistencia a la tracción y son muy estables, con un tipo de cambio lento y poca descomposición, aunque la fosforilación puede promover su desmontaje. Estos son algunos ejemplos populares:
- Las queratinas se encuentran en células epiteliales, células mesodermas y neuronas. Proporcionan fuerza y vienen en formas ácidas y básicas. Cada uno puede formar su propia hebra, pero la mayoría de los IFs consisten en dos hebras, una básica y otra ácida, retorcidas una alrededor de la otra. El cabello y las uñas están hechos de queratina «dura» rica en cisteína para enlaces de disulfuro que proporciona la inmensa fuerza. Las permanentes y el alisado se basan en reducir los enlaces de disulfuro, remodelar el cabello y luego reformar los enlaces de disulfuro. También tiene queratina «suave» en la piel.Los desmanes como la vimentina se encuentran en las células mesenquimales (hueso, cartiledge y grasa).
- Los neurofilamentos se encuentran en axones neuronales y regulan el diámetro de los mismos, lo que a su vez determina la velocidad de propagación potencial de acción.Las láminas
- son las más extendidas y se cree que son más similares al ancestro filogenético de todos los demás IFs. Proporcionan soporte estructural para la membrana nuclear. Podrían ayudar a espaciar los complejos de poros nucleares y también a organizar el ADN.
Finalmente, un video de resumen: