Cellebiologi 07: Mikrotubuli og Celledeling
dette er notater fra forelesning 7 Av Harvard Extensions Cellebiologi kurs.
Forelesning 6 introduserte mikrotubuli, og dette foredraget vil diskutere deres rolle i celledeling. Her er en innledende video:
Oversikt over cellesyklusen
cellesyklusen – prosessen med celledeling og replikering – styres av en rekke biokjemiske brytere kalt cell cycle control system.cellesyklusen er delt inn i faser som er delt inn i faser – folk vil referere til «4 faser», men da er det faktisk 5, og folk bruker også andre ord for å gruppere disse fasene sammen, og andre ord for å dele dem. Jeg har gjort mitt beste for å oppsummere forholdet mellom disse vilkårene i følgende tabell. (modifisert/utvidet Fra Wikipedia):
| MOST general grouping | the supposed «4 phases” | subphases |
|---|---|---|
| non-dividing | Gap 0 (G0) | |
| interphase | Gap 1 (G1) | G1a R G1b |
| Synthesis (S) | ||
| Gap 2 (G2) | ||
| Mitosis | Mitosis (M) | prophase prometaphase metaphase anaphase telophase cytokinesis |
The innholdet i hver fase er vakkert oppsummert I Dette fremragende Wikimedia Commons-bildet Av Kelvinsong:

de raskest delende humane cellene kan fullføre en cellesyklus på omtrent 24 timer (G1: 9h, S: 10h, G2: 4h, M: 30 min). Gjær kan fullføre en syklus på 30 minutter, og de raskest delende Drosophila-cellene tar så lite som 8 minutter.
Hovedkontrollere av denne prosessen inkluderer sykliner, som regulerer syklinavhengig kinase eller CDK. Husk at kinaser er proteiner som fosforylerer andre proteiner. CDKS fosforylering av målene gjør at mitose kan fortsette. For å være presis, er modningsfremmende faktor ELLER MPF et obligatorisk heterodimerisk kompleks bestående av cyklin B og CDK, som bare gjør sin fosforylerende virkning når begge proteiner er tilstede.
Rolle av mikrotubuli
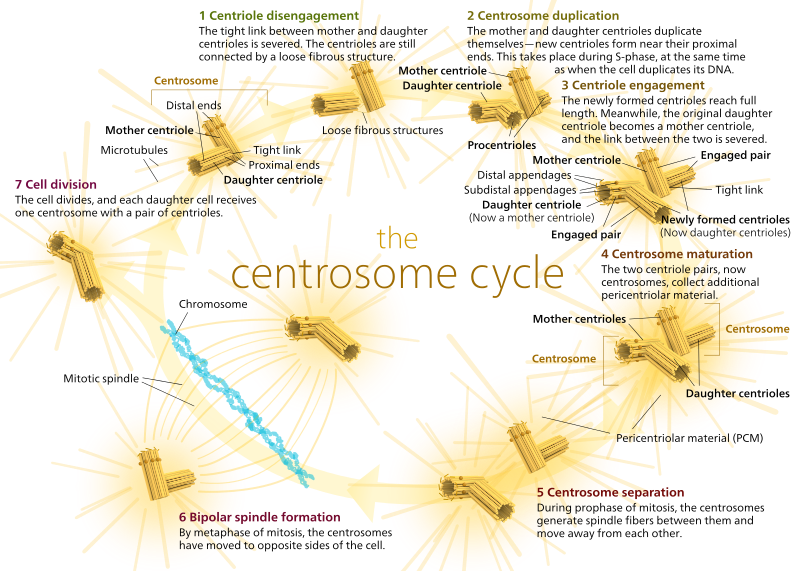
Mikrotubuli er kritiske gjennom cellesyklusen – de organiserer cellulære komponenter og deler dem i to. Her er en serie videoer av cellesyklusen som fremhever rollen som mikrotubuli:hos dyr har hvilende celler og til og med celler i interfase vanligvis bare EN MTOC, kalt et sentrosom, som fungerer som det sentrale navet for alle mikrotubuli i cellen. Et centrosom består av to centrioler som vist nedenfor (takk igjen Til Kelvinsong):

de to sentriolene løsner fra hverandre og replikerer seg selv under s-fasen, og separerer deretter for å danne motsatte «poler» av cellen under m-fasen, slik at det nå er to Mtoc, som hver til slutt vil være den eneste MTOC av en ny celle (et annet boss Kelvinsong-bilde):
under mitose har du de to polene i cellen, hver med mikrotubuli forankret i (-) enden og med deres (+) ender overlappende, peker inn i midten av cellen, som vist her (Wikimedia Commons bilde Av Lordjuppiter):
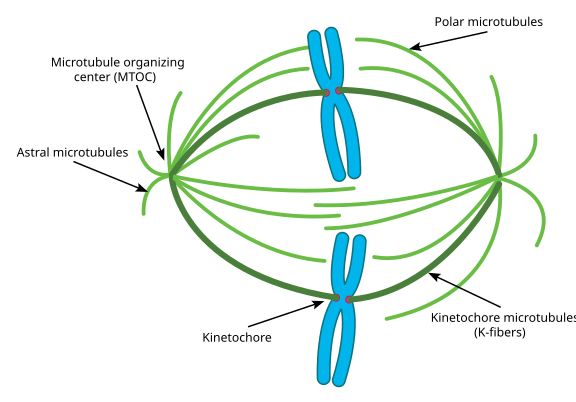
Det hele kalles et spindelapparat, og området der De To MTOCs’ mikrotubuli overlapper kalles ‘ sone av interdigitasjon .’Du vil noen ganger høre HVER MTOC og dens urchin-lignende rekke mikrotubuli kalt en’ mitotisk aster.’
Mikrotubuli under dette stadiet sies å falle inn i tre kategorier:
- Astrale mikrotubuli peker utover, mot cellebarken, for å forankre hele spindelapparatet langs aksen av celledeling.
- Kinetochore mikrotubuli festes til kinetochore av kromatider.Polare mikrotubuli, orientert parallelt med hverandre, men i motsatte retninger, er avgjørende for å skyve spindelapparatet fra hverandre under mitose. (Faktisk er polare mikrotubuli også tilstede tidligere og bidrar til å skyve sentrosomene fra hverandre under profase).
hvis du foretrekker bilder over diagrammer, ser det ut som hele spindelapparatet, med kromatider i blått, mikrotubuli i grønt og kinetochorene som røde prikker:

Sentromere DNA har lav informasjon entropi og spesielle histoner som skiller seg fra andre kromatin. Centromerer er en del av genomet du nesten aldri plukker opp i neste generasjons sekvensering, selv på veldig høy dybde. Det er fordi sentromerer tjener et annet formål enn mye av resten av genomet: sekvensen er gunstig for interaksjon med sentromere proteiner og kinetochore vedlegg. Kohesiner er proteiner som holder de to søsterkromatidene sammen. Vi vil referere til kinetochore proteiner som å ha to lag, den indre kinetochore og ytre kinetochore.
under prometafase beveger kromosomene seg frem og tilbake. Kinesiner forankrer kromosomene til kinetochore mikrotubuli utover spissen der Kinesin-13 depolymeriserer mikrotubuli, hjulpet av mangel på tilgjengelige tubulin dimerer. En kombinasjon av motorproteiner, mikrotubuli-interagerende proteiner og tredemølle tjener til å bevege kromosomene. I mellomtiden, dynein og dynactin – motor proteiner som går mot ( – ) end-arbeid på astrale mikrotubuli, trekke MTOCs mot cellen periferien. I metafase kommer kromatidene til å bli justert langs metafaseplaten.
under denne prosessen oppløses atomkuvertet og slik at atomimport blir irrelevant. Ran-GEF lokaliserer nær kromosomer og genererer høye konsentrasjoner Av Ran-GTP som gir energi til noen nødvendige prosesser (?).
Celler har noen mekanisme for å oppdage spenningen i mikrotubuli som indikerer deres festekromatider før mitose kan fortsette. Å sørge for at hver kromatid er riktig forankret, er avgjørende for å unngå aneuploidi.forresten spiller andre cytoskeletale elementer i tillegg til mikrotubuli også en nøkkelrolle i cellesyklusen. I cytokinesis danner actin en kontraktil ring og, ved hjelp av myosin II motorproteiner, cinches cellen i to.
Betydningen av modellorganismer
oppdagelsen av cellesyklusregulatoriske prosesser stod sterkt på noen fine egenskaper av populære modellorganismer.Saccharomyces cerevisiae (spirende gjær) og Schizosaccharomyces pombe (fisjon gjær) kan eksistere som haploider eller diploider. Det er viktig fordi i haploidfasen kan en mutasjon slå et gen ut – du trenger ikke å treffe begge alleler. Og i gjær er mange mutasjoner, spesielt I Cdc-genene, temperaturavhengige, hvor et protein med en missense-mutasjon fortsatt kan fungere ordentlig ved ‘permissive’ temperaturer, men mister sin opprinnelige funksjon ved’ ikke-permissive ‘ temperaturer. Dette gjør det mulig å studere knockout fenotypen (ved den ikke-permissive temperaturen) mens den fortsatt har det praktiske å kunne forplante organismene (ved den permissive temperaturen). Hele S. cerevisiae-genomet er tilgjengelig som plasmid-biblioteker, noe som gjør det mulig å skjerme for hvilket plasmid redder fenotypen til en gitt mutant. Det er hvor mange av genene som regulerer cellesyklusen ble oppdaget.
I S. cerivisiae, spirende er en del av fase G1, og når dattercellen når en viss størrelse, i et øyeblikk kalt «START», er de to forpliktet Til å gå Inn I S og til slutt fullføre cellesyklusen. Pattedyrceller har sitt eget forpliktelsespunkt kalt begrensningspunktet Eller R, I G1, som er analogt MED START.
Temperaturfølsomme Cdc28-mutanter knopper ikke ved den ikke-tolerante temperaturen. Cdc28-genet koder gjærens homolog av VÅR syklinavhengig kinase (CDK) som, når og bare når kompleks med syklin, kan fosforylere andre proteiner for å regulere deres deltakelse i cellesyklusfaser. Temperaturfølsomme mutanter ved nonpermissive temperaturen sitter fast i stand til å knopp og gå Inn I s-fasen. I stedet oppfører de seg som villtype celler berøvet næringsstoffer: de vokser store nok til å passere START, men fortsetter ikke.
Xenopus (en slags frosk) viste seg kritisk for å forstå cellesyklus, fordi reproduksjonen innebærer et meget stort antall celler (dvs . nok utgangsmateriale For Western blots, etc.) som er perfekt synkronisert (dvs. alle er i samme fase av cellesyklusen i samme øyeblikk. (Sammenlign med gjær, for eksempel, hvor celler ikke alle vil være i samme fase samtidig). Også egget selv er stort og lett å jobbe med, og flere cellesykluser følger befruktning. I frosker begynner egg meiotisk deling, men arresterer deretter I g2-fasen i 8 måneder mens de vokser i størrelse og lager ting som vil være nødvendig for vekst ved befruktning.I tillegg til mikrofilamenter og mikrotubuli har eukaryote celler også en rekke andre cytoskeletale proteiner kalt intermediære filamenter (ifs). Selv om det er mer variert enn mikrofilamenter og mikrotubuli, Er IFs Ikke bare et fangstbegrep for ‘ noe annet filament – – heller, de er en gruppe relaterte proteiner. De strekker seg vanligvis gjennom cytoplasma og indre kjernehylse, er ikke-polare og har ingen motorproteiner forbundet med dem. De har stor strekkstyrke og er veldig stabile, med en langsom valutakurs og ikke mye sammenbrudd, selv om fosforylering kan fremme demontering. Her er noen populære eksempler:
- Keratiner finnes i epitelceller, mesodermceller og nevroner. De gir styrke og kommer i sure og grunnleggende former. Hver kan danne sin egen tråd, men de fleste If består av to tråder-en grunnleggende og en sur, slags vridd rundt hverandre. Hår og negler er laget av ‘hard’ keratin rik på cystein for disulfidbindinger som gir den enorme styrken. Perm og rette stole på å redusere disulfidbindingene, omforme håret og deretter reformere disulfidbindingene. Du har også ‘myk’ keratin i huden din.
- Desmins som vimentin finnes i mesenkymale celler(bein, kartiledge og fett).
- Nevrofilamenter er i nevronaksoner og regulere diameteren derav, som i sin tur bestemmer hastigheten på aksjonspotensial forplantning.
- Laminer er begge de mest utbredte Og antas å være mest lik fylogenetisk stamfar til alle de andre IFs. De gir strukturell støtte til kjernemembranen. De kan hjelpe plass ut de kjernefysiske porekompleksene og også organisere DNA.
Endelig en oppsummering video: