«Hvor lang tid tar de forskjellige stadiene av cellesyklusen?
Hvor lang tid tar de forskjellige stadiene av cellesyklusen?
Lesermodus
Replikering er en av kjennetegnene til levende materie. Settet av prosesser kjent som cellesyklusen som gjennomføres når en celle blir to, har vært et dominerende forskningstema i molekylærtiden med applikasjoner som strekker seg langt og bredt, inkludert til studiet av sykdommer som kreft som noen ganger karakteriseres som en sykdom i cellesyklusen gått galt. Cellesykluser er interessante både for måtene de ligner fra en celletype til den neste og for måtene de er forskjellige på. For å bringe emnet i lettelse, vurderer vi cellesyklusene i en rekke forskjellige organismer, inkludert en modellprokaryote, for pattedyrceller i vevskultur og under embryonisk utvikling i fruktfluen. Spesielt spør vi hva er de enkelte trinnene som gjennomføres for en celle å dele seg i to og hvor lenge tar disse trinnene?
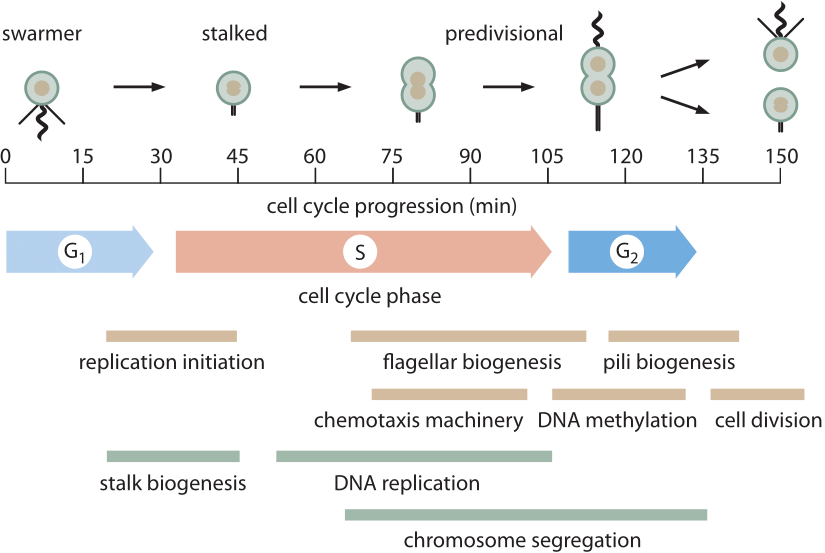
Figur 1: 150 min celle syklus Av Caulobacter er vist, fremhever noen av de viktigste morfologiske og metabolske hendelser som finner sted under celledeling. M-fase er ikke indikert fordi I Caulobacter er det ikke noe sant mitotisk apparat som blir montert som i eukaryoter. Mye av kromosomsegregering i Caulobacter (og andre bakterier) skjer samtidig MED DNA-replikasjon. De siste trinnene av kromosomsegregering og spesielt dekatenering av de to sirkulære kromosomene oppstår under G2-fasen. (Tilpasset Fra M. T. Laub et al., Vitenskap 290:2144, 2000.)
Uten Tvil den best karakteriserte prokaryote cellesyklusen er den av modellorganismen Caulobacter crescentus. En av de tiltalende egenskapene til denne bakterien er at den har en asymmetrisk celledeling som gjør det mulig for forskere å binde en av de to avkomene til et mikroskopdeksel mens den andre datteren driver bort slik at videre studier uten hindringer. Dette har gitt grundige avbildninger av den ≈150 minutters cellesyklusen (BNID 104921) som vist I Figur 1. Hovedkomponentene i cellesyklusen Er G1 (første Vekstfase, ≈30 min, BNID 104922), hvor minst en minimal mengde cellestørrelsesøkning må finne sted, S-fase (Syntese, ≈80 min, BNID 104923) HVOR DNA blir replikert og G2 (andre Vekstfase, ≈25 min, BNID 104924) hvor kromosomsegregering utfolder seg som fører til celledeling (sluttfase varig ≈15 min). Caulobacter crescentus gir et interessant eksempel på hvordan visse organismer blir forfremmet til» modellorganisme » – status fordi de har en spesiell funksjon som gjør dem spesielt passende for spørsmålet om interesse. I dette tilfellet går cellesyklusprogresjonen hånd i hånd med differensieringsprosessen som gir lett visualiserte identifiserbare stadier, noe som gjør dem foretrukket for cellesyklusbiologer over, sier modellbakterien E. coli.oppførselen til pattedyrceller i vevskultur har tjent som grunnlag for mye av det vi vet om cellesyklusen i høyere eukaryoter. Den eukaryotiske cellesyklusen kan grovt deles inn i to trinn, interfase, den delen av cellesyklusen når materialene i cellen blir duplisert og mitose, settet av fysiske prosesser som deltar i kromosomsegregering og påfølgende celledeling. Satsene for prosesser i cellesyklusen, er for det meste bygget opp fra mange av de molekylære hendelsene som polymerisering AV DNA og cytoskeletal filamenter hvis priser vi allerede har vurdert. For den karakteristiske cellesyklustiden på 20 timer i En HeLa-celle er nesten halvparten viet Til G1 (BNID 108483) og nær en annen halvdel Er S-fase (BNID 108485) mens G2 og M er mye raskere på henholdsvis 2-3 timer og 1 time (bnid 109225, 109226). Scenen mest variabel i varighet Er G1. I mindre gunstige vekstforhold når cellesyklusvarigheten øker, er dette scenen som for det meste påvirkes, sannsynligvis på grunn av tiden det tar før noen regulatorisk størrelse sjekkpunkt er nådd. Selv om ulike typer bevis peker på eksistensen av et slikt kontrollpunkt, er det for tiden svært dårlig forstått. Historisk har stadier i cellesyklusen vanligvis blitt utledet ved hjelp av faste celler, men nylig har genetisk kodede biosensorer som endrer lokalisering på forskjellige stadier av cellesyklusen gjort det mulig å få live-celle temporal informasjon om cellesyklusprogresjon og arrestasjon.
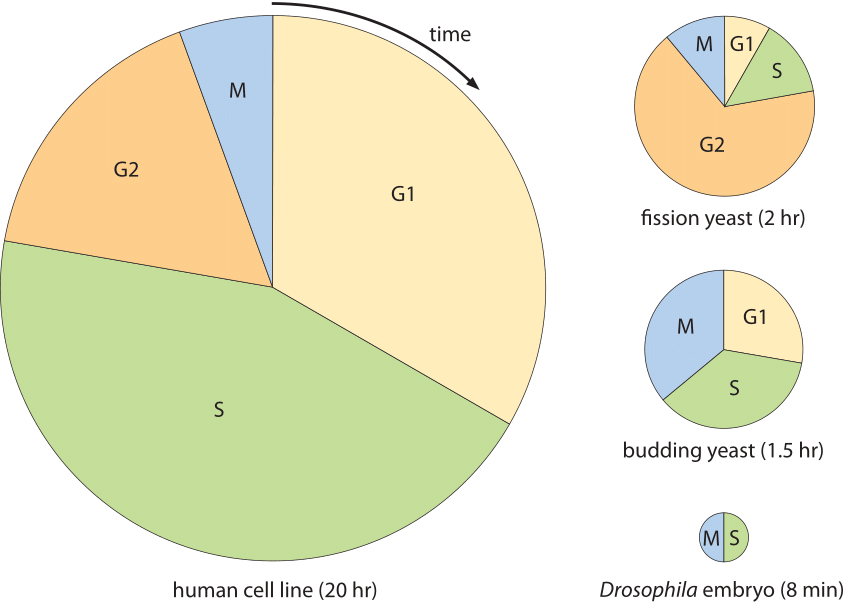
Figur 2: Cellesyklustider for forskjellige celletyper. Hvert kakediagram viser brøkdelen av cellesyklusen viet til hvert av de primære stadiene av cellesyklusen. Arealet av hvert diagram er proporsjonalt med den totale cellesyklusvarigheten. Cellesyklusvarigheter reflekterer minimale doblingstider under ideelle forhold. (Tilpasset fra» The Cell Cycle – Principles Of Control » Av David Morgan.)
hvordan sammenligner lengden på cellesyklusen med tiden det tar en celle å syntetisere sitt nye genom? En avkobling mellom genomlengden og doblingstiden eksisterer i eukaryoter på grunn av bruken av FLERE DNA-replikasjonsstartsteder. For pattedyrceller har det blitt observert at for mange vev med svært varierende totale cellesyklustider, er varigheten av s-fasen der DNA-replikasjon oppstår bemerkelsesverdig konstant. For musevev som de som finnes i tykktarmen eller tungen, varierte s-fasen i et lite område fra 6,9 til 7,5 timer (BNID 111491). Selv når man sammenligner flere epitelvev på tvers av mennesker, rotter, mus og hamster, var S-fasen mellom 6 og 8 timer (BNID 107375). Disse målingene ble utført på 1960 – tallet ved å utføre en slags pulsjakteksperiment med radioaktivt merket nukleotidtymidin. Under den korte puls ble den radioaktive forbindelsen bare innlemmet i genomet av celler I s-fase. Ved å måle varigheten av utseende og deretter forsvinning av merkede celler I m fase kan man utlede hvor lenge s fase varte det faktum at varigheten Av s fase er relativt konstant i slike celler brukes til denne dag for å estimere varigheten av cellesyklusen fra en kunnskap om bare brøkdel av celler på et gitt øyeblikksbilde i tid som er I s fase. For eksempel, hvis en tredjedel av cellene er sett I S-fase som varer ca 7 timer, er cellesyklustiden utledes å være ca 7 timer/(1/3) ≈20 timer. I dag er disse typer målinger er for det meste utført Ved Hjelp Av BrdU som markør For s fase. Vi er ikke klar over en tilfredsstillende forklaring på opprinnelsen til denne relativt konstante replikasjonstiden og hvordan den er relatert TIL frekvensen AV DNA-polymerase og tettheten av replikeringsinitieringssteder langs genomet.
mangfoldet av cellesykluser er vist I Figur 2 og skildrer flere modellorganismer og varigheten og plasseringen av de forskjellige stadiene av deres cellesykluser. Et ekstremt eksempel oppstår i den fascinerende prosessen med embryonisk utvikling av fruktfluen Drosophila melanogaster. I dette tilfellet er situasjonen forskjellig fra konvensjonelle celledivisjoner siden i stedet for å syntetisere nye cytoplasmatiske materialer, er masse i hovedsak bevart bortsett fra replikering av det genetiske materialet. Dette skjer på en veldig synkron måte i ca 10 generasjoner, og en replikasjonssyklus av tusenvis av celler i embryoet, si mellom syklus 10 og 11, skjer i ca 8 minutter som vist i Figur 2 (BNID 103004, 103005, 110370). Dette er raskere enn replikasjonstidene for noen bakterier, selv om genomet er ≈120 millioner bp langt (BNID 100199). Et slående eksempel på cellens evne til å tilpasse sin tidsmessige dynamikk.




