Konsepter Av Biologi-1st Canadian Edition
Virus ble først oppdaget etter utviklingen av et porselenfilter, kalt Chamberland-Pasteur-filteret, som kunne fjerne alle bakterier som er synlige under mikroskopet fra en hvilken som helst væskeprøve. I 1886 viste Adolph Meyer at en sykdom av tobakksplanter, tobaksmosaikksykdom, kunne overføres fra en syk plante til en sunn gjennom flytende planteekstrakter. I 1892 Viste Dmitri Ivanowski at denne sykdommen kunne overføres på denne måten selv etter At Chamberland-Pasteur-filteret hadde fjernet alle levedyktige bakterier fra ekstraktet. Likevel var det mange år før det ble bevist at disse «filtrerbare» smittsomme stoffene ikke bare var svært små bakterier, men var en ny type liten, sykdomsfremkallende partikkel.
Virioner, enkeltviruspartikler, er svært små, ca 20-250 nanometer (1 nanometer = 1/1 000 000 mm). Disse individuelle viruspartiklene er den smittsomme formen av et virus utenfor vertscellen. I motsetning til bakterier (som er omtrent 100 ganger større), kan vi ikke se virus med et lysmikroskop, med unntak av noen store virioner av poxvirusfamilien (Figur 12.3).
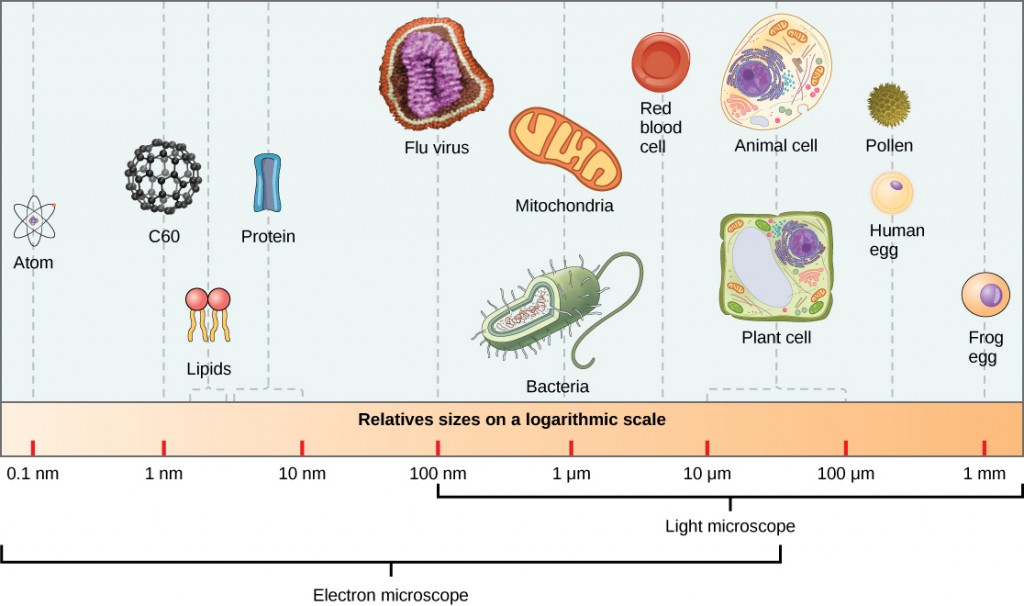
det var ikke før utviklingen av elektronmikroskopet på 1940-tallet at forskerne fikk sitt første gode syn på strukturen av tobaksmosaikkviruset (Figur 12.2) Og andre. Overflatestrukturen av virioner kan observeres ved både skanning og transmisjonselektronmikroskopi, mens virusets indre strukturer kun kan observeres i bilder fra et transmisjonselektronmikroskop (Figur 12.4).
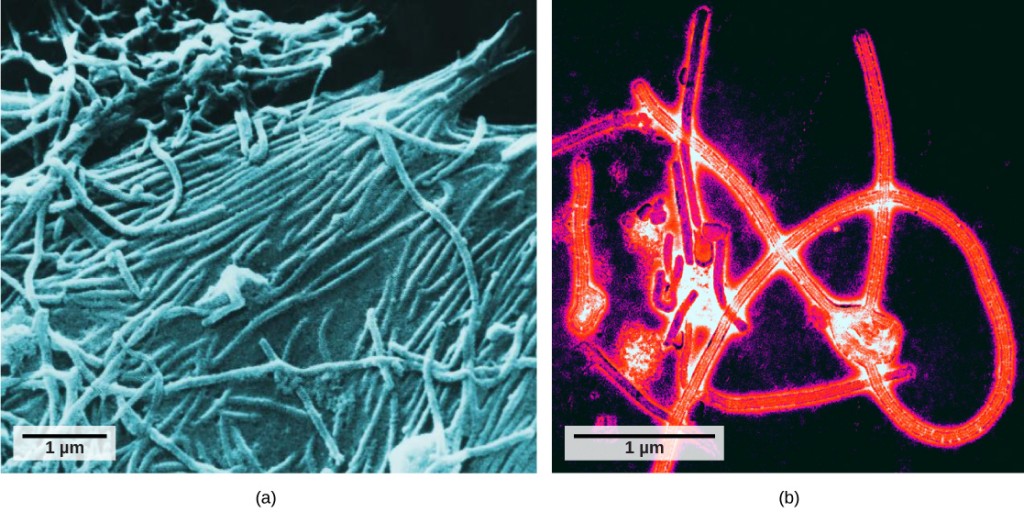
bruken av denne teknologien har gjort det mulig for oppdagelsen av mange virus av alle typer levende organismer. De ble opprinnelig gruppert etter felles morfologi, som betyr deres størrelse, form og skille strukturer. Senere ble grupper av virus klassifisert etter typen nukleinsyre de inneholdt, DNA ELLER RNA, og om deres nukleinsyre var enkelt-eller dobbeltstrenget. Mer nylig har molekylær analyse av virusreplikasjonssykluser ytterligere raffinert klassifiseringen.en virion består av en nukleinsyrekjerne, et ytre proteinbelegg, og noen ganger en ytre konvolutt laget av protein og fosfolipidmembraner avledet fra vertscellen. Den mest synlige forskjellen mellom medlemmer av virale familier er deres morfologi, som er ganske variert. Et interessant trekk ved viral kompleksitet er at kompleksiteten til verten ikke korrelerer med kompleksiteten til virionen. Noen av de mest komplekse virionstrukturer observeres i bakteriofager, virus som smitter de enkleste levende organismer, bakterier.
Virus kommer i mange former og størrelser, men Disse er konsistente og forskjellige for hver virusfamilie (Figur 12.5). Alle virioner har et nukleinsyregenom dekket av et beskyttende lag av protein, kalt en kapsid. Kapsid er laget av protein subenheter kalt capsomeres. Noen virale kapsler er enkle polyhedrale «sfærer», mens andre er ganske komplekse i struktur. Den ytre strukturen rundt kapsid av noen virus kalles viral konvolutt. Alle virus bruker en slags glykoprotein for å feste seg til vertscellene ved molekyler på cellen som kalles virale reseptorer. Viruset utnytter disse celleoverflatemolekylene, som cellen bruker til et annet formål, som en måte å gjenkjenne og infisere bestemte celletyper. For eksempel bruker meslingviruset et celleoverflateglykoprotein hos mennesker som normalt fungerer i immunreaksjoner og muligens i sperm-egg-interaksjonen ved befruktning. Vedlegg er et krav for at virus senere skal trenge inn i cellemembranen, injisere virusgenomet og fullføre replikasjonen inne i cellen.T4-bakteriofagen, som infiserer e. coli-bakterien, er blant de mest komplekse virionene som er kjent; T4 har en proteinhalestruktur som viruset bruker til å feste til vertscellen og en hodestruktur som huser SITT DNA.Adenovirus, et nonenveloped dyrevirus som forårsaker respiratoriske sykdommer hos mennesker, bruker proteinpigger som stikker ut fra sine kapsomerer for å feste seg til vertscellen. Nonenveloped virus inkluderer også de som forårsaker polio (poliovirus), plantarvorter (papillomavirus) og hepatitt a (hepatitt a-virus). Nonenveloped virus har en tendens til å være mer robust og mer sannsynlig å overleve under tøffe forhold, for eksempel tarmen.Innhyllet virioner SOM HIV (humant immunsviktvirus), det forårsakende middel I AIDS (ervervet immunsvikt syndrom), består av nukleinsyre (RNA I TILFELLE AV HIV) og kapsid proteiner omgitt av en fosfolipid dobbeltlag konvolutt og tilhørende proteiner (Figur 12.5). Vannkopper, influensa og kusma er eksempler på sykdommer forårsaket av virus med konvolutter. På grunn av skjørheten i konvolutten, nonenveloped virus er mer motstandsdyktig mot endringer i temperatur, pH, og noen desinfeksjonsmidler enn innhyllet virus.Samlet sett forteller formen på virionen og tilstedeværelsen eller fraværet av en konvolutt oss lite om hvilke sykdommer virusene kan forårsake eller hvilke arter de kan infisere, Men er fortsatt et nyttig middel for å begynne viral klassifisering.
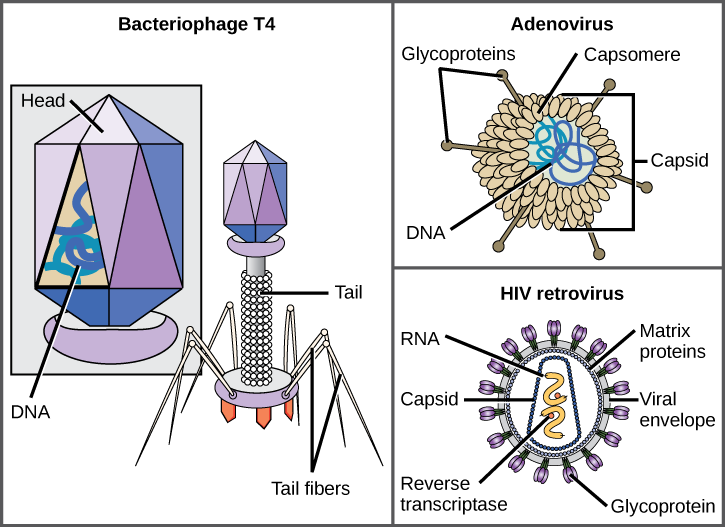
hvilke av følgende utsagn om virusstruktur er sanne?
A) alle virus er innkapslet i en virusmembran.
B) capsomeren består av små proteinunderenheter kalt capsids.
C) DNA ER det genetiske materialet i alle virus.
D) Glykoproteiner hjelper viruset å feste seg til vertscellen.
<!–D->
I Motsetning til alle levende organismer som bruker DNA som sitt genetiske materiale, kan virus bruke ENTEN DNA eller RNA som deres. Viruskjernen inneholder genomet eller det totale genetiske innholdet av viruset. Virale genomer har en tendens til å være små sammenlignet med bakterier eller eukaryoter, som bare inneholder de genene som koder for proteiner viruset ikke kan få fra vertscellen. Dette genetiske materialet kan være enkeltstrenget eller dobbeltstrenget. Det kan også være lineært eller sirkulært. Mens de fleste virus inneholder et enkelt segment av nukleinsyre, har andre genomer som består av flere segmenter.
DNA-virus har EN DNA-kjerne. Det virale DNA styrer vertscellens replikasjonsproteiner for å syntetisere nye kopier av det virale genomet og å transkribere og oversette det genomet til virale proteiner. DNA-virus forårsaker menneskelige sykdommer som vannkopper, hepatitt B og noen veneriske sykdommer som herpes og kjønnsvorter.
RNA-virus inneholder bare RNA i deres kjerner. For å replikere deres genomer i vertscellen, koder genomene AV RNA-virus enzymer som ikke finnes i vertsceller. RNA-polymerase-enzymer er ikke like stabile SOM DNA-polymeraser og gjør ofte feil under transkripsjon. Av denne grunn forekommer mutasjoner, endringer i nukleotidsekvensen, I RNA-virus oftere enn I DNA-virus. Dette fører til raskere utvikling og endring I RNA-virus. For eksempel er det faktum at influensa ER ET RNA-virus en grunn til at en ny influensavaksine er nødvendig hvert år. Menneskelige sykdommer forårsaket AV RNA-virus inkluderer hepatitt C, meslinger og rabies.
Virus kan ses som obligatoriske intracellulære parasitter. Viruset må feste seg til en levende celle, tas inn, produsere proteiner og kopiere genomet, og finne en måte å unnslippe cellen slik at viruset kan infisere andre celler og til slutt andre individer. Virus kan infisere bare visse arter av verter og bare visse celler i den verten. Det molekylære grunnlaget for denne spesifisiteten er at et bestemt overflatemolekyl, kjent som viral reseptor, må finnes på vertscellens overflate for at viruset skal feste seg. Også metabolske forskjeller sett i forskjellige celletyper basert på differensial genuttrykk er en sannsynlig faktor der celler et virus kan bruke til å replikere. Cellen må lage stoffene viruset trenger, for eksempel enzymer virusgenomet selv ikke har gener for, eller viruset vil ikke kunne replikere ved hjelp av den cellen.
Trinn Av Virusinfeksjoner
et virus må «ta over» en celle for å replikere. Den virale replikasjonssyklusen kan produsere dramatiske biokjemiske og strukturelle endringer i vertscellen, noe som kan forårsake celleskader. Disse endringene, kalt cytopatiske effekter, kan endre cellefunksjoner eller til og med ødelegge cellen. Noen infiserte celler, som de som er smittet av forkjølelsesvirus (rhinovirus), dør gjennom lysis (sprengning) eller apoptose(programmert celledød eller «celle selvmord»), og frigjør alle avkomvirionene samtidig. Symptomene på virussykdommer skyldes immunresponsen mot viruset, som forsøker å kontrollere og eliminere viruset fra kroppen, og fra celleskader forårsaket av viruset. Mange dyrevirus, SOM HIV (humant immunsviktvirus), forlater de infiserte cellene i immunsystemet ved en prosess kjent som spirende, hvor virioner forlater cellen individuelt. Under den spirende prosessen gjennomgår cellen ikke lys og blir ikke umiddelbart drept. Imidlertid kan skaden på cellene SOM HIV smitter gjøre det umulig for cellene å fungere som mediatorer av immunitet, selv om cellene forblir i live i en periode. Mest produktive virusinfeksjoner følger lignende trinn i virusreplikasjonssyklusen: vedlegg, penetrasjon, ubestrøket, replikering, montering og utgivelse.Et virus festes til et bestemt reseptorsted på vertscellemembranen gjennom festeproteiner i kapsidet eller proteiner innebygd i konvolutten. Vedlegget er spesifikt, og vanligvis vil et virus bare feste seg til celler av en eller noen få arter og bare visse celletyper innenfor de artene med de riktige reseptorene.
Konsept i Aksjon

Se denne videoen for en visuell forklaring på HVORDAN HIV og influensa angriper kroppen.
I Motsetning til dyrevirus injiseres nukleinsyren av bakteriofager i vertscellen naken, og forlater kapsidet utenfor cellen. Plante-og dyrevirus kan komme inn i sine celler gjennom endocytose, hvor cellemembranen omgir og engulfs hele viruset. Noen innhyllede virus kommer inn i cellen når viruskuvertet smelter direkte med cellemembranen. Når du er inne i cellen, blir den virale kapsid degradert og den virale nukleinsyren frigjøres, som deretter blir tilgjengelig for replikasjon og transkripsjon.
replikasjonsmekanismen avhenger av det virale genomet. DNA-virus bruker vanligvis vertscelleproteiner og enzymer for å lage ekstra DNA som brukes til å kopiere genomet eller transkriberes til messenger RNA (mRNA), som deretter brukes i proteinsyntese. RNA-virus, som influensaviruset, bruker vanligvis rna-kjernen som en mal for syntese av viral genomisk RNA og mRNA. Den virale mRNA blir oversatt til virale enzymer og kapsidproteiner for å samle nye virioner (Figur 12.6). Selvfølgelig er det unntak fra dette mønsteret. Hvis en vertscelle ikke gir enzymer som er nødvendige for viral replikasjon, gir virale gener informasjonen til direkte syntese av de manglende proteinene. Retrovirus, SOM HIV, har ET RNA-genom som må reverseres transkribert FOR Å lage DNA, som deretter settes inn i vertsens DNA. For å konvertere RNA TIL DNA inneholder retrovirus gener som koder for det virusspesifikke enzymet revers transkriptase som transkriberer EN RNA-mal TIL DNA. DET FAKTUM AT HIV produserer noen av sine egne enzymer, som ikke finnes i verten, har gjort det mulig for forskere å utvikle stoffer som hemmer disse enzymene. Disse stoffene, inkludert revers transkriptaseinhibitoren AZT, hemmer HIV-replikasjon ved å redusere enzymets aktivitet uten å påvirke vertens metabolisme.
den siste fasen av viral replikasjon er frigjøringen av de nye virionene i vertsorganismen, hvor de er i stand til å infisere tilstøtende celler og gjenta replikasjonssyklusen. Noen virus frigjøres når vertscellen dør og andre virus kan forlate infiserte celler ved å spire gjennom membranen uten å drepe cellen direkte.
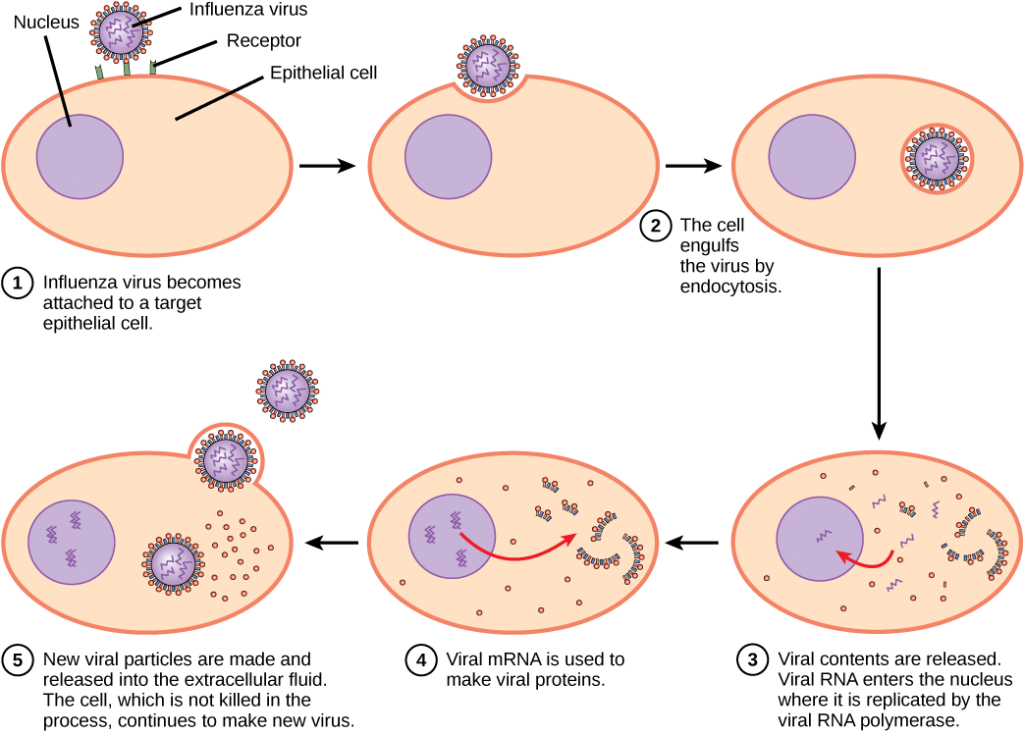
Influensavirus er pakket i en viral konvolutt som smelter sammen med plasmamembranen. På denne måten kan viruset gå ut av vertscellen uten å drepe det. Hvilken fordel får viruset ved å holde vertscellen i live?
<!- Vertscellen kan fortsette å lage nye viruspartikler.– >
Konsept I Aksjon

Klikk gjennom denne opplæringen om virus for å identifisere strukturer, overføringsmåter, replikering og mer.