Biologia komórki 07: mikrotubule i podział komórek
Oto notatki z wykładu 7 kursu biologii komórki Harvard Extension.
Wykład 6 przedstawił mikrotubule, a ten wykład omówi ich rolę w podziale komórek. Oto film wprowadzający:
przegląd cyklu komórkowego
cykl komórkowy – proces podziału i replikacji komórek – jest regulowany przez serię biochemicznych przełączników zwanych systemem kontroli cyklu komórkowego.
cykl komórkowy jest podzielony na fazy, które są podzielone na fazy – ludzie będą odnosić się do „4 faz”, ale w rzeczywistości jest ich 5, a ludzie używają również innych słów do grupowania tych faz razem, a inne słowa do ich podziału. Dołożyłem wszelkich starań, aby podsumować związek między tymi Warunkami w poniższej tabeli. (zmodyfikowane / rozbudowane z Wikipedii):
| MOST general grouping | the supposed „4 phases” | subphases |
|---|---|---|
| non-dividing | Gap 0 (G0) | |
| interphase | Gap 1 (G1) | G1a R G1b |
| Synthesis (S) | ||
| Gap 2 (G2) | ||
| Mitosis | Mitosis (M) | prophase prometaphase metaphase anaphase telophase cytokinesis |
The zawartość każdej fazy jest pięknie podsumowana na tym wyjątkowym obrazku Wikimedia Commons autorstwa Kelvinsonga:

najszybciej dzielące się komórki ludzkie mogą ukończyć cykl komórkowy w około 24 godziny (G1: 9h, S: 10h, G2: 4h, M: 30 min). Drożdże mogą zakończyć cykl w 30 minut, a najszybciej dzielące się komórki Drosophila zajmują zaledwie 8 minut.
główne Kontrolery tego procesu obejmują cykliny, które regulują kinazę zależną od cyklin lub CDK. Przypomnijmy, że kinazy są białkami, które fosforylują inne białka. Fosforylacja CDK umożliwia mitozę. Aby być precyzyjnym, czynnik promujący dojrzewanie lub MPF jest zobowiązującym kompleksem heterodimerycznym złożonym z cykliny B i CDK, który działa fosforylująco tylko wtedy, gdy oba białka są obecne.
rola mikrotubul
mikrotubule mają kluczowe znaczenie w całym cyklu komórkowym – organizują komponenty komórkowe i dzielą je na dwie części. Oto seria filmów z cyklu komórkowego, które podkreślają rolę mikrotubul:
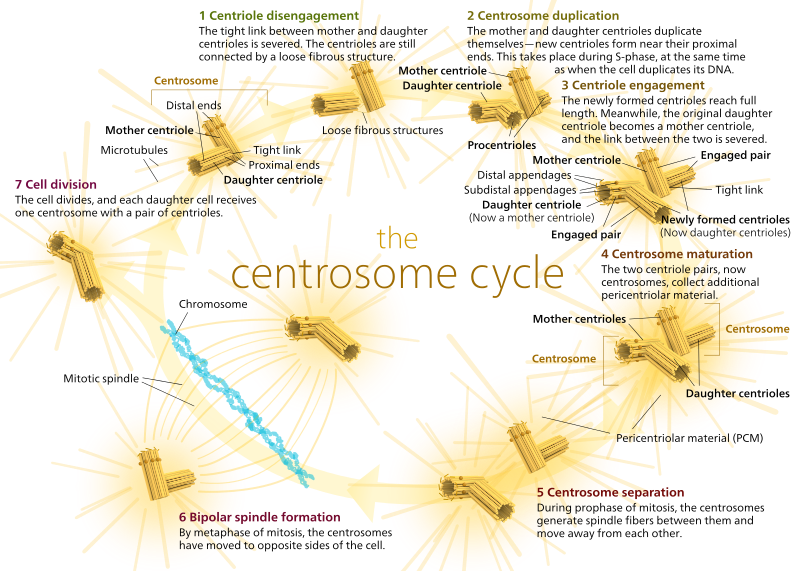
u zwierząt komórki spoczynkowe, a nawet komórki międzyfazowe, zwykle mają tylko jeden MTOC, zwany centrosomem, który służy jako centralny hub dla wszystkich mikrotubul w komórce. Centrosom składa się z dwóch centrioli, jak pokazano poniżej (jeszcze raz dzięki Kelvinsongowi):

dwa centriole odłączają się od siebie i replikują się podczas fazy S, a następnie oddzielają się, tworząc przeciwległe”bieguny”komórki podczas fazy M, tak że teraz są dwa Mtoc, z których każdy ostatecznie będzie jedynym MTOC nowej komórki (inny obraz kelvinsonga bossa):
w trakcie mitozy mamy dwa „bieguny” komórki, każdy z mikrotubulami zakotwiczonymi na końcu ( – ) i z ich końcami ( + ) nakładającymi się na środek komórki, jak pokazano tutaj (obraz Wikimedia Commons autorstwa Lordjuppitera):
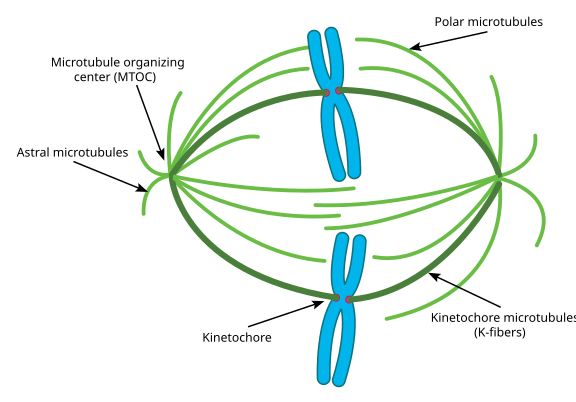
to wszystko nazywa się przyrządem wrzecionowatym, a obszar, w którym dwie mikrotubule MTOCs nakładają się na siebie, nazywa się ” strefą interdygacji.’Czasami usłyszysz każdy MTOC i jego podobny do jeżowca zestaw mikrotubul zwany’ asterem mitotycznym.”
mikrotubule na tym etapie dzielą się na trzy kategorie:
- mikrotubule Astralne wskazują na zewnątrz, w kierunku kory komórkowej, aby zakotwiczyć cały aparat wrzeciona wzdłuż osi podziału komórki.
- mikrotubule Kinetochorowe przyłączają się do kinetochoru chromatyd.
- mikrotubule Polarne, zorientowane równolegle do siebie, ale w przeciwnych kierunkach, są kluczowe dla rozsunięcia aparatu wrzeciona podczas mitozy. (W rzeczywistości mikrotubule polarne są również obecne wcześniej i pomagają odepchnąć centrosomy podczas profazy).
jeśli wolisz zdjęcia zamiast diagramów, oto jak wygląda cały aparat wrzeciona, z chromatydami na niebiesko, mikrotubulami na zielono, a kinetochorami jako czerwonymi kropkami:

mikrotubule stają się znacznie bardziej dynamiczne podczas mitozy: więcej gamma-tubuliny sprzyja łatwiejszemu zarodkowaniu, ale xmap215, stabilizator mikrotubul, jest fosforylowany i w ten sposób inaktywowany podczas mitozy, pozostawiając kinezynę-13 wolną do katastrofy mikrotubul. Fortuny szybko się zbijają i tracą. Okres półtrwania mikrotubuli podczas mitozy wynosi około 15 minut, w porównaniu do 30 minut podczas interfazy. Ludzie badają dynamikę mikrotubuli za pomocą FRAP: dodaj fluorescencyjną mikrotubulę, wybiel ją i zobacz, jak szybko następuje ponowny montaż w oparciu o to, jak szybko pojawia się fluorescencja. + Wskazówki odgrywają również ważną rolę w wspomaganiu i montażu mikrotubul.
Kinezyna-5 ma dwie głowy biegunowe, które wiążą się z przeciwstawnymi mikrotubulami i próbują iść w kierunku (+) końca każdej z nich. To odpycha dwie mikrotubule od siebie i zapewnia siłę napędową do oddzielenia MTOCs.
Centromeryczne DNA ma niską entropię informacyjną i specjalne histony, które różnią się od innych chromatyny. Centromery są częścią genomu, którego prawie nigdy nie wykrywa się w sekwencjonowaniu nowej generacji, nawet na bardzo dużej głębokości. To dlatego, że centromery służą innym celom niż większość reszty genomu: Sekwencja tam jest korzystna dla interakcji z białkami centromerycznymi i przyłączeniem kinetochoru. Kohezyny są białkami, które utrzymują razem dwie siostrzane chromatydy. Będziemy odnosić się do białek kinetochorowych jako mających dwie warstwy, kinetochor wewnętrzny i kinetochor zewnętrzny.
podczas prometafazy chromosomy poruszają się tam iz powrotem. Kinezyny zakotwiczają chromosomy w mikrotubulach kinetochorowych poza końcówką, gdzie Kinezyna – 13 depolimeryzuje mikrotubule, wspomagane przez niedobór dostępnych dimerów tubuliny. Połączenie białek motorycznych, białek oddziałujących na mikrotubule i treadmilling służy do przemieszczania chromosomów. Tymczasem dyneina i dynaktyna – białka motoryczne, które podążają w kierunku ( – ) końca-pracują na mikrotubulach Astralnych, ciągnąc MTOCs w kierunku obrzeża komórki. W metafazie chromatydy są ustawione wzdłuż „płytki metafazy”.
podczas tego procesu powłoka jądrowa rozpuszcza się, a więc import jądrowy staje się nieistotny. Ran-GEF lokalizuje się w pobliżu chromosomów i generuje wysokie stężenia Ran-GTP, które dostarcza energii dla niektórych niezbędnych procesów (?).
komórki mają pewien mechanizm wykrywania napięcia w mikrotubulach, który wskazuje na ich przyłączenie przed rozpoczęciem mitozy. Upewnienie się, że każda chromatyda jest prawidłowo zakotwiczona, jest kluczowe dla uniknięcia aneuploidii.
przy okazji, inne elementy cytoszkieletu oprócz mikrotubul również odgrywają kluczową rolę w cyklu komórkowym. W cytokinezie aktyna tworzy pierścień kurczliwy i przy pomocy białek motorycznych miozyny II cynkuje komórkę na dwie części.
Znaczenie organizmów modelowych
odkrycie procesów regulacji cyklu komórkowego w dużym stopniu opierało się na pewnych cechach popularnych organizmów modelowych.
Saccharomyces cerevisiae (drożdże budujące) i Schizosaccharomyces pombe (drożdże rozszczepialne) mogą występować jako haploidy lub diploidy. To ważne, ponieważ w fazie haploidalnej jedna mutacja może wybić gen – nie trzeba uderzać w oba allele. A w drożdżach wiele mutacji, zwłaszcza w genach Cdc _ _ (Kontrola podziału komórek), zależy od temperatury, gdzie białko z mutacją missense może nadal funkcjonować prawidłowo w temperaturach „permisywnych”, ale traci swoją natywną funkcję w temperaturach „nie permisywnych”. Umożliwia to badanie fenotypu knockout (w temperaturze nie-permisywnej), a jednocześnie ma wygodę bycia w stanie łatwo rozmnażać organizmy (w temperaturze permisywnej). Cały genom S. cerevisiae jest dostępny jako biblioteki plazmidów, dzięki czemu możliwe jest prześwietlenie, dla którego plazmid ratuje fenotyp danego mutanta. Tak wiele genów regulujących cykl komórkowy zostało odkrytych.
W S. cerivisiae, pączkowanie jest częścią fazy G1, a gdy komórka potomna osiągnie pewien rozmiar, w momencie zwanym „startem”, obie zobowiązują się do wejścia w S i ostatecznie zakończenia cyklu komórkowego. Komórki ssaków mają swój własny punkt zaangażowania zwany punktem ograniczenia lub R, W G1, który jest analogiczny do początku.
wrażliwe na temperaturę mutanty Cdc28 nie budzą się w temperaturze nietrwałej. Gen Cdc28 koduje homolog drożdży z naszej kinazy zależnej od cyklin (CDK), która, kiedy i tylko wtedy, gdy jest skompleksowana z cykliną, może fosforylować inne białka, aby regulować ich udział w fazach cyklu komórkowego. Wrażliwe na temperaturę mutanty w temperaturze nietrwałej utkną w stanie pączkowania i wejdą w fazę S. Zamiast tego zachowują się jak komórki typu dzikiego pozbawione składników odżywczych: rosną wystarczająco duże, aby przejść START, ale potem nie kontynuują.
Xenopus (rodzaj żaby) okazał się krytyczny dla zrozumienia cyklu komórkowego, ponieważ jego rozmnażanie obejmuje bardzo dużą liczbę komórek (tj. wystarczająco dużo materiału wyjściowego dla zachodnich plam itp.), które są doskonale zsynchronizowane (tzn. wszystkie znajdują się w tej samej fazie cyklu komórkowego w tym samym momencie. (Porównaj na przykład z drożdżami, gdzie komórki nie będą wszystkie w tej samej fazie w tym samym czasie). Również samo jajo jest duże i łatwe w obsłudze, a po zapłodnieniu następuje wiele cykli komórkowych. U żab jaja rozpoczynają podział mejotyczny, a następnie zatrzymują się w fazie G2 przez 8 miesięcy, podczas gdy rosną i gromadzą rzeczy potrzebne do wzrostu po zapłodnieniu.
włókna pośrednie
oprócz mikrofilamentów i mikrotubul, komórki eukariotyczne mają również wiele „innych” białek cytoszkieletowych zwanych włóknami pośrednimi (IFs). Chociaż są bardziej zróżnicowane niż mikrofilamenty i mikrotubule, IFs nie są tylko chwytliwym terminem dla „każdego innego włókna” – są raczej grupą pokrewnych białek. Na ogół rozciągają się przez cytoplazmę i wewnętrzną otoczkę jądrową, są niepolarne i nie mają z nimi powiązanych białek motorycznych. Mają dużą wytrzymałość na rozciąganie i są bardzo stabilne, z powolnym kursem wymiany i niewielkim rozpadem, chociaż fosforylacja może promować ich demontaż. Oto kilka popularnych przykładów:
- keratyny znajdują się w komórkach nabłonkowych, komórkach mezodermy i neuronach. Zapewniają siłę i występują w postaci kwaśnej i zasadowej. Każda może tworzyć własne nici, ale większość IFs składa się z dwóch nici-jednej zasadowej i jednej kwaśnej, jakby skręconych wokół siebie. Włosy i paznokcie wykonane są z „twardej” keratyny bogatej w cysteinę dla wiązań dwusiarczkowych, która zapewnia ogromną siłę. Trwałe i prostowanie polegają na zmniejszeniu wiązań dwusiarczkowych, przekształcaniu włosów, a następnie reformowaniu wiązań dwusiarczkowych. Masz również „miękką” keratynę w skórze.
- Desminy takie jak wimentyna znajdują się w komórkach mezenchymalnych (kości, kości i tkanki tłuszczowej).
- Neurofilamenty znajdują się w aksonach neuronalnych i regulują ich średnicę, co z kolei determinuje szybkość propagacji potencjału czynnościowego.
- laminy są najbardziej rozpowszechnione i uważa się, że są najbardziej podobne do filogenetycznego przodka wszystkich pozostałych Ifów. Zapewniają one wsparcie strukturalne dla błony jądrowej. Mogą pomóc w rozmieszczeniu atomowych kompleksów porów, a także zorganizować DNA.
wreszcie filmik podsumowujący: