Cell Biology 07: microtúbulos and Cell Division
These are notes from lecture 7 of Harvard Extension’s Cell Biology course.
Lecture 6 introduziu microtúbulos, e esta palestra irá discutir o seu papel na divisão celular. Aqui está um vídeo introdutório:
Visão geral do ciclo celular
O ciclo celular – o processo de divisão celular e a replicação é regido por uma série de parâmetros bioquímicos chamado o sistema de controle do ciclo celular.
O Ciclo Celular é dividido em fases que são divididas em fases – as pessoas se referirão às “4 fases”, mas então há realmente 5, e as pessoas também usam outras palavras para agrupar essas fases juntas, e outras palavras para subdividi-las. Fiz o meu melhor para resumir a relação entre estes termos na seguinte tabela. (modificado/expandido da Wikipédia):
| MOST general grouping | the supposed “4 phases” | subphases |
|---|---|---|
| non-dividing | Gap 0 (G0) | |
| interphase | Gap 1 (G1) | G1a R G1b |
| Synthesis (S) | ||
| Gap 2 (G2) | ||
| Mitosis | Mitosis (M) | prophase prometaphase metaphase anaphase telophase cytokinesis |
The o conteúdo de cada fase é bem resumida em destaque no Wikimedia Commons imagem de Kelvinsong:

O mais rápido-divisão de células humanas pode completar um ciclo celular em cerca de 24 horas (G1: 9h, S: 10h, G2: 4h, M: 30 min). A levedura pode terminar um ciclo em 30 minutos, e as células de Drosófila que se dividem mais rapidamente levam apenas 8 minutos.os controladores principais deste processo incluem as ciclinas, que regulam a cinase dependente de ciclinas ou CDK. Lembre-se que as cinases são proteínas que fosforilato outras proteínas. A fosforilação dos alvos do CDK permite que a mitose prossiga. Para ser preciso, o Fator de maturação ou MPF é um complexo heterodimérico obrigatório composto de cyclin B E CDK, que só faz sua ação de fosforilação quando ambas as proteínas estão presentes.os microtúbulos são críticos ao longo do ciclo celular – organizam componentes celulares e dividem-nos em dois. Aqui estão uma série de vídeos do ciclo celular que destacam o papel dos microtúbulos:
em animais, células Quiescentes e até mesmo células em interfase geralmente têm apenas um MTOC, chamado de centrossoma, que serve como centro central para todos os microtúbulos na célula. Um centrosome é composto por dois centrióis como mostrado abaixo (Graças novamente a Kelvinsong):

Os dois centrioles soltar um do outro e se reproduzem durante a fase S, e depois se separam para formar em frente ‘dos pólos da célula durante o M de fase, de modo que agora existem dois MTOCs, cada um dos quais, eventualmente, será o único MTOC de uma nova célula (outro chefe Kelvinsong imagem):
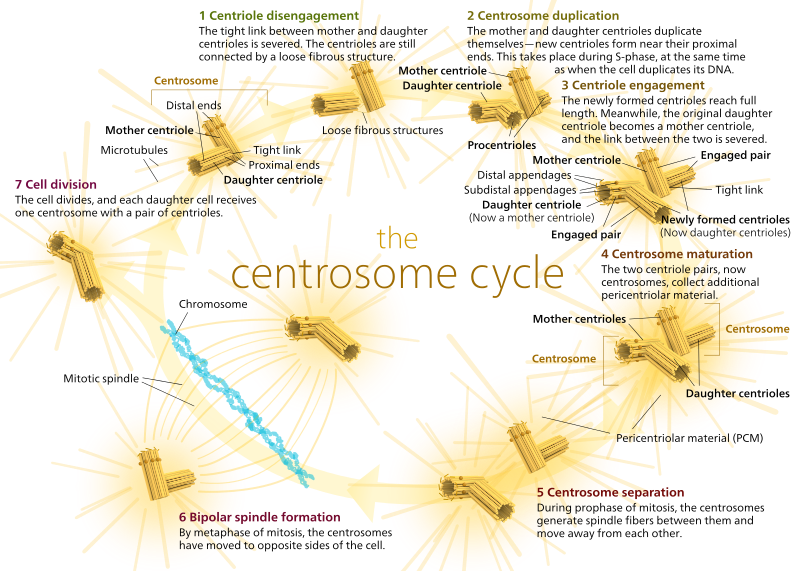
Durante a mitose, em seguida, você tem os dois “pólos” da célula, cada um com microtúbulos ancorado no (-) final e com a sua (+) extremidades sobrepostas, apontando para o centro da célula, como mostrado aqui (Wikimedia Commons imagem de Lordjuppiter):
essa coisa toda é chamada de um aparato de fuso, e a área onde os microtúbulos dos dois MTOCs se sobrepõem é chamada de “zona de interdigitação”. Às vezes você vai ouvir cada MTOC e seu conjunto de microtúbulos chamado de “aster mitótico”.os microtúbulos ”
durante esta fase são classificados em três categorias:
- microtúbulos Astrais apontam para fora, em direcção ao córtex celular, a fim de ancorar todo o aparato do fuso ao longo do eixo da divisão celular.microtúbulos Cinetócoros ligam-se ao cinetócoro dos cromatídeos.microtúbulos polares, orientados paralelamente uns aos outros, mas em direções opostas, são cruciais para separar o aparato de fuso durante a mitose. (Na verdade, microtúbulos polares também estão presentes mais cedo e ajudam a separar os centrossomas durante a profase).
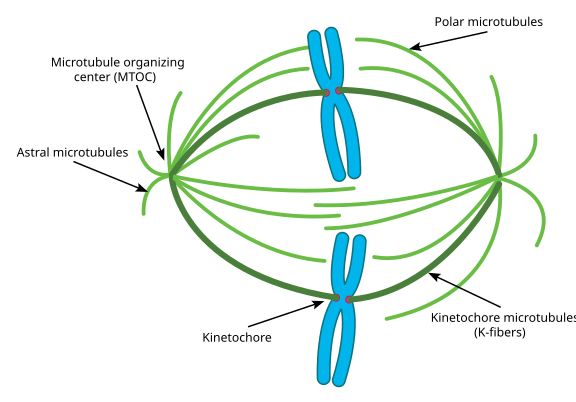
Se preferir fotos em vez de diagramas, aqui está o aspecto de todo o aparato de fuso, com cromatídeos em azul, microtúbulos em verde, e os cinetóforos como pontos vermelhos:

microtúbulos tornam-se muito mais dinâmicos durante a mitose: mais Gama-tubulina promove nucleação mais fácil, mas XMAP215, um estabilizador de microtúbulos, é fosforilado e, portanto, inactivado durante a mitose, deixando a Cinesin-13 livre para catastrofizar os microtúbulos. Fortunas são feitas e perdidas rapidamente. A semi-vida de um microtúbulo durante a mitose é de cerca de 15 minutos, em comparação com 30 minutos durante a interfase. As pessoas estudam a dinâmica de microtúbulos usando FRAP: adicione um microtúbulo fluorescente, descolore-o e veja quão rápido a remontagem está ocorrendo com base em quão breve a fluorescência reaparece. + Dicas também desempenham um papel importante na ajuda e montagem dos microtúbulos.
Cinesin-5 Is tem duas cabeças polares que se ligam a microtúbulos opostos e tentam caminhar em direção ao fim (+) de cada uma. Isto afasta os dois microtúbulos e fornece a força motriz para a separação dos Mtoc.o ADN Centromérico tem pouca informação sobre entropia e histonas especiais que diferem de outra cromatina. Centrômeros são uma parte do genoma que quase nunca se apanha em sequenciação de próxima geração, mesmo a uma profundidade muito alta. Isso porque os centrômeros estão servindo a um propósito diferente de grande parte do resto do genoma: a sequência lá é favorável para a interação com proteínas centrômicas e apego cinetócoro. As coesinas são proteínas que mantêm os dois cromatídeos Irmãos Unidos. Vamos referir-nos às proteínas cinetocore como tendo duas camadas, a cinetocore interior e a cinetocore exterior.durante a prometafase, os cromossomas movem-se para trás e para a frente. As cinesinas ancoram os cromossomas nos microtúbulos cinetócoros para além da ponta, onde a cinesina-13 despolimeriza os microtúbulos, auxiliada pela escassez de dimeros de tubulina disponíveis. Uma combinação de proteínas motoras, proteínas interagindo microtúbulos e treadmilling serve para mover os cromossomos. Entretanto, as proteínas motoras da dineína e da dinactina que caminham em direcção ao ( – ) trabalho final nos microtúbulos Astrais, puxando os MTOCs para a periferia celular. Em metafase, os cromatídeos são alinhados ao longo da “placa de metafase”.
durante este processo, o envelope nuclear dissolve-se e, por isso, a importação nuclear torna-se irrelevante. Ran-GEF localiza perto dos cromossomas e gera altas concentrações de Ran-GTP que fornece energia para alguns processos necessários (?).as células
têm algum mecanismo para detectar a tensão nos microtúbulos que indica os seus cromatídeos de ligação antes de a mitose poder prosseguir. Certificar-se de que cada cromatídeo está devidamente ancorado é crucial para evitar a aneuploidia.a propósito, outros elementos do citoesqueleto além dos microtúbulos também desempenham um papel fundamental no ciclo celular. Na citoquinese, a actina forma um anel contractil e, com a ajuda das proteínas motoras da miosina II, cinta a célula em duas.
importância dos organismos modelo
A descoberta de processos regulatórios do ciclo celular dependia fortemente de algumas características de organismos modelo populares.Saccharomyces cerevisiae (levedura) e Schizosaccharomyces pombe (levedura de fissão) podem existir sob a forma de haploides ou diplóides. Isso é importante porque na fase haploid, uma mutação pode derrubar um gene – você não precisa acertar em ambos os alelos. E na levedura, muitas mutações, especialmente no Cdc__ (controle de divisão celular) genes, são dependentes da temperatura, onde uma proteína com uma mutação missense ainda pode funcionar corretamente a temperaturas “permissivas”, mas perde sua função nativa a temperaturas “não permissivas”. Isto torna possível estudar o fenótipo knockout (à temperatura não permissiva) enquanto ainda tendo a conveniência de ser capaz de propagar facilmente os organismos (à temperatura permissiva). Todo o genoma da S. cerevisiae está disponível como bibliotecas plasmídicas, tornando possível rastrear para que plasmídeo resgata o fenótipo de um determinado mutante. É o número de genes que regulam o ciclo celular que foram descobertos.em S. cerivisiae, budding é parte da fase G1, e uma vez que a célula filha atinge um certo tamanho, em um momento chamado “início”, os dois estão comprometidos em entrar S e, finalmente, completar o ciclo celular. As células de mamíferos têm seu próprio ponto de compromisso chamado de ponto de restrição ou R, em G1, O que é análogo ao início.os mutantes Cdc28 sensíveis à temperatura não brotam à temperatura não permissiva. O gene Cdc28 codifica o homolog da levedura da nossa cinase dependente de ciclinas (CDK) que, quando e somente quando complexada com a ciclina, pode fosforilar outras proteínas para regular a sua participação nas fases do ciclo celular. Mutantes sensíveis à temperatura à temperatura não permissiva ficam presos incapazes de brotar e entrar na fase S. Em vez disso, eles se comportam como células do tipo selvagem privadas de nutrientes: eles crescem grande o suficiente para passar início, mas, em seguida, não continuar.Xenopus (um tipo de rã) provou ser crítico para a compreensão do ciclo celular, pois sua reprodução envolve um grande número de células. material de partida suficiente para blots ocidentais, etc.) que estão perfeitamente sincronizados (ou seja, todos estão na mesma fase do ciclo celular no mesmo momento. (Compare com a levedura, por exemplo, onde as células não estarão todas na mesma fase ao mesmo tempo). Também o ovo em si é grande e fácil de trabalhar, e múltiplos ciclos celulares seguem a fertilização. Em sapos, os ovos começam a divisão meiótica, mas em seguida, prender na fase G2 por 8 meses, enquanto eles crescem em tamanho e armazenar coisas que serão necessárias para o crescimento após a fertilização.além dos microfilamentos e microtúbulos, as células eucarióticas têm também uma série de outras proteínas citoesqueléticas denominadas filamentos intermédios (IFs). Embora mais diversificados do que microfilamentos e microtúbulos, os IFs não são apenas um termo geral para “qualquer outro filamento”, mas sim um grupo de proteínas relacionadas. Eles geralmente se estendem através do citoplasma e envelope nuclear interno, são não-polares e não têm proteínas motoras associadas a eles. Eles têm grande resistência à tracção e são muito estáveis, com uma taxa de câmbio lenta e não muito colapso, embora a fosforilação pode promover a sua desmontagem. Aqui estão alguns exemplos populares:
- queratinas são encontradas em células epiteliais, células mesodérmicas e neurônios. Eles fornecem força e vêm em formas ácidas e básicas. Cada um pode formar a sua própria cadeia, mas a maioria dos Fi consistem em duas cadeias – uma básica e uma ácida, Tipo torcida em torno da outra. O cabelo e as unhas são feitos de queratina “dura” rica em cisteína para ligações de dissulfeto que fornece a imensa força. Os Perms e o alisamento dependem da redução das ligações de dissulfeto, remodelando o cabelo e, em seguida, reformando as ligações de dissulfeto. Você também tem queratina ‘suave’ em sua pele.Desminas como a vimentina são encontradas em células mesenquimais (osso, cartiledge e gordura).os Neurofilamentos encontram-se em axônios neuronais e regulam o seu diâmetro, o que, por sua vez, determina a velocidade de propagação potencial de ação.
- laminas são os mais difundidos e acredita-se que sejam mais semelhantes ao ancestral filogenético de todos os outros IFs. Eles fornecem suporte estrutural para a membrana nuclear. Eles podem ajudar a criar espaço para os complexos de poros nucleares e também organizar o DNA.
finalmente, um vídeo sumário: