Biologie celulară 07: microtubuli și diviziune celulară
acestea sunt note din cursul 7 al cursului de Biologie Celulară al extensiei Harvard.
prelegerea 6 a introdus microtubuli, iar această prelegere va discuta despre rolul lor în diviziunea celulară. Iată un videoclip introductiv:
Prezentare generală a ciclului celular
ciclul celular – procesul de diviziune și replicare celulară – este guvernat de o serie de comutatoare biochimice numite sistemul de control al ciclului celular.
ciclul celular este împărțit în faze care sunt împărțite în faze – oamenii se vor referi la „4 faze”, dar apoi există de fapt 5, iar oamenii folosesc și alte cuvinte pentru a grupa aceste faze și alte cuvinte pentru a le subdiviza. Am făcut tot posibilul pentru a rezuma relația dintre acești termeni în tabelul următor. (modificat/extins de pe Wikipedia):
| MOST general grouping | the supposed „4 phases” | subphases |
|---|---|---|
| non-dividing | Gap 0 (G0) | |
| interphase | Gap 1 (G1) | G1a R G1b |
| Synthesis (S) | ||
| Gap 2 (G2) | ||
| Mitosis | Mitosis (M) | prophase prometaphase metaphase anaphase telophase cytokinesis |
The conținutul fiecărei faze este frumos rezumat în această imagine remarcabilă Wikimedia Commons de Kelvinsong:

celulele umane cu cea mai rapidă divizare pot finaliza un ciclu celular în aproximativ 24 de ore (G1: 9h, S: 10h, G2: 4h, M: 30 min). Drojdia poate termina un ciclu în 30 de minute, iar celulele Drosophila cu cea mai rapidă divizare durează doar 8 minute.controlerele principale ale acestui proces includ ciclinele, care reglează kinaza dependentă de ciclină sau CDK. Amintiți-vă că kinazele sunt proteine care fosforilează alte proteine. Fosforilarea țintelor sale de către CDK permite mitozei să continue. Pentru a fi mai precis, factorul de promovare a maturării sau MPF este un complex heterodimeric obligatoriu compus din ciclina B și CDK, care își face acțiunea de fosforilare numai atunci când sunt prezente ambele proteine.
rolul microtubulilor
microtubulii sunt critici pe tot parcursul ciclului celular – organizează componentele celulare și le împart în două. Iată o serie de videoclipuri ale ciclului celular care evidențiază rolul microtubulilor:
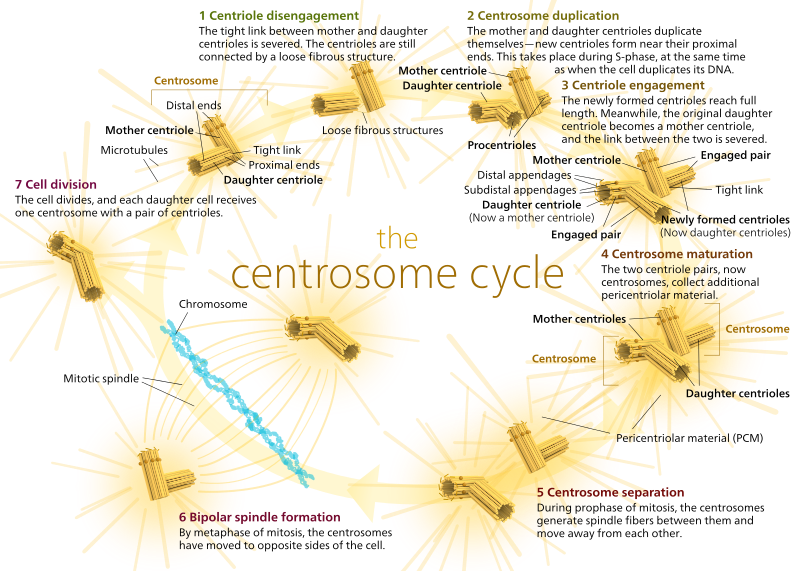
la animale, celulele în repaus și chiar celulele din interfază au de obicei un singur MTOC, numit centrosom, care servește ca hub central pentru toți microtubulii din celulă. Un centrosom este format din doi centrioli așa cum se arată mai jos (datorită din nou Kelvinsong):

cei doi centrioli se desprind unul de celălalt și se reproduc în timpul fazei S, apoi se separă pentru a forma”poli”opuși ai celulei în timpul fazei M, astfel încât acum există două Mtoc-uri, Fiecare dintre acestea va fi în cele din urmă singurul MTOC al unei celule noi (o altă imagine Boss Kelvinsong):
în timpul mitozei, atunci, aveți cei doi”poli”ai celulei, fiecare cu microtubuli ancorați la capătul (-) și cu capetele lor (+) suprapuse, îndreptate în centrul celulei, așa cum se arată aici (imaginea Wikimedia Commons de Lordjuppiter):
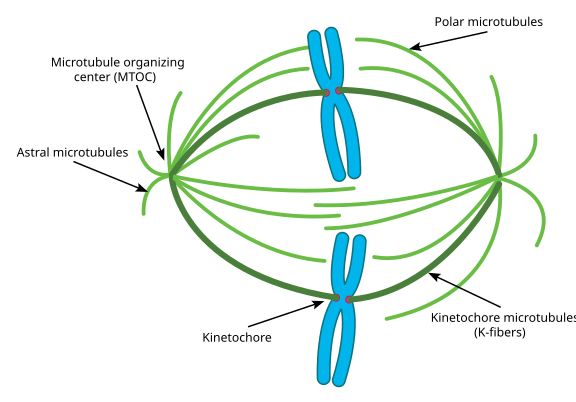
Acest lucru este numit un aparat de ax, și zona în care cele două MTOCs’ microtubuli se suprapun este numit ‘zona de interdigitație. Veți auzi uneori fiecare MTOC și gama sa de microtubuli asemănători urchinului numită Aster mitotic.’
microtubulii în această etapă se spune că se încadrează în trei categorii:
- microtubulii astrali indică spre exterior, spre cortexul celular, pentru a ancora întregul aparat al fusului de-a lungul axei diviziunii celulare.
- microtubulii Kinetochor se atașează la kinetochorul cromatidelor.
- microtubulii polari, orientați paralel unul cu celălalt, dar în direcții opuse, sunt cruciali pentru împingerea aparatului axului în afară în timpul mitozei. (De fapt, microtubulii polari sunt prezenți și mai devreme și ajută la împingerea centrozomilor în timpul profazei).
dacă preferați fotografiile în locul diagramelor, iată cum arată întregul aparat cu ax, cu cromatide în albastru, microtubuli în verde și kinetochori ca puncte roșii:

microtubulii devin mult mai dinamici în timpul mitozei: mai mult gamma-tubulină promovează nucleația mai ușoară, dar XMAP215, un stabilizator de microtubuli, este fosforilat și astfel inactivat în timpul mitozei, lăsând Kinesina-13 liberă să catastrofeze microtubulii. Averile sunt făcute și pierdute rapid. Timpul de înjumătățire al unui microtubul în timpul mitozei este de aproximativ 15 minute, comparativ cu 30 de minute în timpul interfazei. Oamenii studiază dinamica microtubulilor folosind FRAP: adăugați un microtubul fluorescent, înălbiți-l și vedeți cât de repede se produce reasamblarea în funcție de cât de curând reapare fluorescența. + Sfaturile joacă, de asemenea, un rol major în sprijinirea și asamblarea microtubulilor.
Kinesin-5 is are două capete polare care se leagă de microtubuli opuși și încearcă să meargă spre capătul (+) al fiecăruia. Aceasta împinge cei doi microtubuli în afară și asigură forța motrice pentru separarea Mtoc-urilor.
ADN-ul Centromeric are Entropie informațională scăzută și histone speciale care diferă de alte cromatine. Centromerii sunt o parte a genomului pe care aproape niciodată nu o detectați în secvențierea următoarei generații, chiar și la o adâncime foarte mare. Acest lucru se datorează faptului că centromerii servesc unui scop diferit de o mare parte din restul genomului: secvența de acolo este favorabilă interacțiunii cu proteinele centromerice și atașamentul kinetocor. Coezinele sunt proteine care păstrează cele două cromatide surori împreună. Ne vom referi la proteinele kinetochore ca având două straturi, kinetochorul interior și kinetochorul exterior.
în timpul prometafazei, cromozomii se mișcă înainte și înapoi. Kinesinele ancorează cromozomii la microtubulii kinetocori dincolo de vârful în care Kinesina-13 depolimerizează microtubulii, ajutată de o lipsă de dimeri tubulinici disponibili. O combinație de proteine motorii, proteine care interacționează cu microtubuli și banda de alergare servește la mișcarea cromozomilor. Între timp, dineina și dynactina – proteinele motorii care merg spre (-) end – lucrează la microtubulii astrali, trăgând Mtoc-urile spre periferia celulei. În metafază, cromatidele ajung să fie aliniate de-a lungul’plăcii metafazei’.
în timpul acestui proces, învelișul nuclear se dizolvă și astfel importul nuclear devine irelevant. Ran-GEF localizează în apropierea cromozomilor și generează concentrații mari de Ran-GTP care furnizează energie pentru unele procese necesare (?).
celulele au un mecanism pentru detectarea tensiunii în microtubuli care indică cromatidele lor de atașament înainte ca mitoza să poată continua. Asigurarea faptului că fiecare cromatidă este ancorată corespunzător este crucială pentru evitarea aneuploidiei.apropo, alte elemente citoscheletice în afară de microtubuli joacă, de asemenea, un rol cheie în ciclul celular. În citokineză, actina formează un inel contractil și, cu ajutorul proteinelor motorii miozinei II, cinchează celula în două.
importanța organismelor model
descoperirea proceselor de reglare a ciclului celular s-a bazat foarte mult pe unele caracteristici îngrijite ale organismelor model populare.Saccharomyces cerevisiae (drojdie în devenire) și Schizosaccharomyces pombe (drojdie de fisiune) pot exista ca haploizi sau diploizi. Acest lucru este important, deoarece în faza haploidă, o mutație poate elimina o genă – nu trebuie să lovești ambele alele. Și în drojdie, multe mutații, în special în genele CDC (controlul diviziunii celulare), sunt dependente de temperatură, unde o proteină cu o mutație missense poate funcționa în continuare corect la temperaturi ‘permisive’, dar își pierde funcția nativă la temperaturi ‘nepermisive’. Acest lucru face posibilă studierea fenotipului knockout (la temperatura nepermisivă), având în același timp comoditatea de a putea propaga cu ușurință organismele (la temperatura permisivă). Întregul genom S. cerevisiae este disponibil ca biblioteci de plasmide, făcând posibilă examinarea pentru care plasmida salvează fenotipul unui mutant dat. Așa au fost descoperite multe dintre genele care reglează ciclul celular.
în S. cerivisiae, înmugurirea face parte din faza G1 și, odată ce celula fiică atinge o anumită dimensiune, într-un moment numit „START”, cei doi se angajează să intre în S și, în cele din urmă, să finalizeze ciclul celular. Celulele mamiferelor au propriul punct de angajament numit punctul de restricție sau R, în G1, care este analog cu START.
mutanții Cdc28 sensibili la temperatură nu mugesc la temperatura nepermisivă. Gena Cdc28 codifică omologul drojdiei kinazei noastre dependente de ciclină (CDK) care, atunci când și numai atunci când este complexată cu ciclină, poate fosforila alte proteine pentru a-și regla participarea la fazele ciclului celular. Mutanții sensibili la temperatură la temperatura nepermisivă se blochează incapabili să mugească și să intre în faza S. În schimb, se comportă ca celulele de tip sălbatic lipsite de nutrienți: cresc suficient de mari pentru a trece de la început, dar apoi nu continuă.
Xenopus (un fel de broască) s-a dovedit critic pentru înțelegerea ciclului celular, deoarece reproducerea sa implică un număr foarte mare de celule (adică. material de pornire suficient pentru bloturile occidentale etc.) care sunt perfect sincronizate (adică toate sunt în aceeași fază a ciclului celular în același moment. (Comparați cu drojdia, de exemplu, unde celulele nu vor fi toate în aceeași fază în același timp). De asemenea, oul în sine este mare și ușor de lucrat, iar ciclurile celulare multiple urmează fertilizarea. La broaște, ouăle încep diviziunea meiotică, dar apoi se opresc la faza G2 timp de 8 luni, în timp ce cresc în dimensiune și stochează lucruri care vor fi necesare pentru creștere la fertilizare.
filamente intermediare
În plus față de microfilamente și microtubuli, celulele eucariote au, de asemenea, o serie de alte proteine citoscheletice numite filamente intermediare (if). Deși sunt mai diverse decât microfilamentele și microtubulii, if-urile nu sunt doar un termen general pentru ‘orice alt filament’ – mai degrabă, ele sunt un grup de proteine înrudite. În general, se extind prin citoplasmă și învelișul nuclear interior, sunt nepolare și nu au proteine motorii asociate cu acestea. Ei au o mare rezistență la tracțiune și sunt foarte stabile, cu un curs de schimb lent și nu de mult defalcare, deși fosforilarea poate promova demontarea lor. Iată câteva exemple populare:
- keratinele se găsesc în celulele epiteliale, celulele mezoderm și neuronii. Ele oferă rezistență și vin în forme acide și de bază. Fiecare își poate forma propriul fir, dar majoritatea if – urilor constau din două fire-una bazică și una acidă, fel de răsucite unul în jurul celuilalt. Parul si unghiile sunt facute din keratina’ tare ‘ bogata in cisteina pentru legaturi disulfidice care ofera puterea imensa. Permanentul și îndreptarea se bazează pe reducerea legăturilor disulfidice, remodelarea părului și apoi reformarea legăturilor disulfidice. De asemenea, aveți keratină moale în piele.
- Desminele precum vimentina se găsesc în celulele mezenchimale (os, cartiledge și grăsime).
- Neurofilamentele sunt în axoni neuronali și reglează diametrul acestora, care la rândul său determină viteza de propagare a potențialului de acțiune.Laminele sunt ambele cele mai răspândite și se crede că sunt cele mai asemănătoare cu strămoșul filogenetic al tuturor celorlalte if. Ele oferă suport structural pentru membrana nucleară. Ei ar putea ajuta spațiu în complexele porilor nucleare și, de asemenea, organiza ADN-ul.
în cele din urmă, un videoclip rezumat: