Concepts of Biology-1st Canadian Edition
virukset löydettiin ensimmäisen kerran Chamberland-Pasteur-suodattimeksi kutsutun posliinisuodattimen kehittämisen jälkeen, joka pystyi poistamaan kaikki mikroskoopilla näkyvät bakteerit mistä tahansa nestemäisestä näytteestä. Vuonna 1886 Adolph Meyer osoitti, että tupakkakasvien tauti, tupakkamosaiikkitauti, voitiin siirtää sairaasta kasvista terveeseen nestemäisten kasviuutteiden kautta. Vuonna 1892 Dmitri Ivanowski osoitti, että tauti voisi tarttua tällä tavalla senkin jälkeen, kun Chamberland-Pasteur-suodatin oli poistanut kaikki elinkykyiset bakteerit uutteesta. Kului kuitenkin monta vuotta, ennen kuin todistettiin, että nämä ”saastuttavat” taudinaiheuttajat eivät olleet vain hyvin pieniä bakteereja, vaan uudentyyppisiä pieniä, tauteja aiheuttavia hiukkasia.
virionit, yksittäiset viruspartikkelit, ovat hyvin pieniä, noin 20-250 nanometriä (1 nanometri = 1/1 000 000 mm). Nämä yksittäiset viruspartikkelit ovat isäntäsolun ulkopuolisen viruksen tarttuva muoto. Toisin kuin bakteerit (jotka ovat noin 100 kertaa suurempia), emme voi nähdä viruksia valomikroskoopilla, lukuun ottamatta joitakin suuria rokkovirusten suvun virioita (Kuva 12.3).
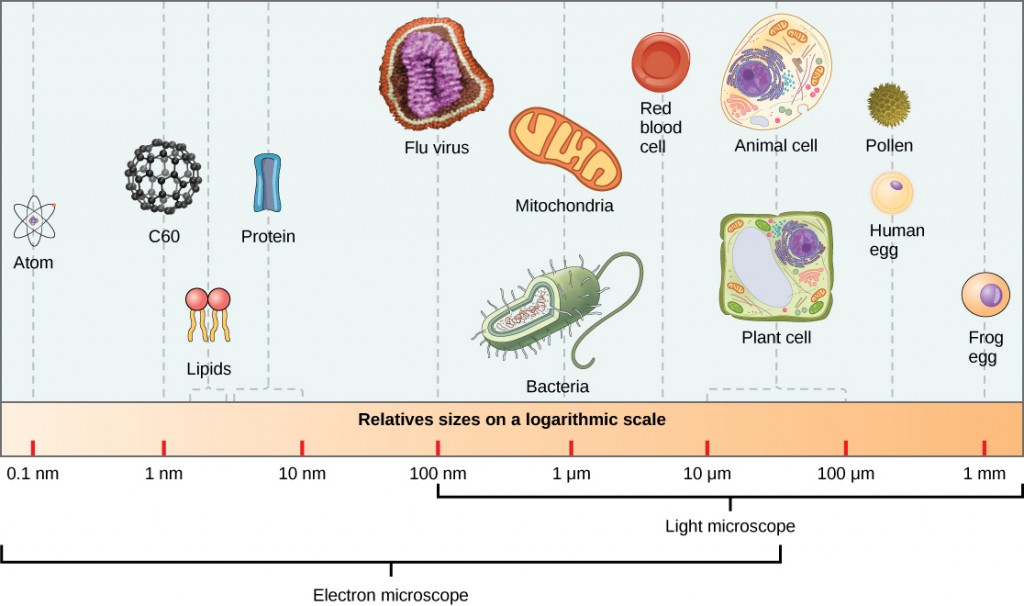
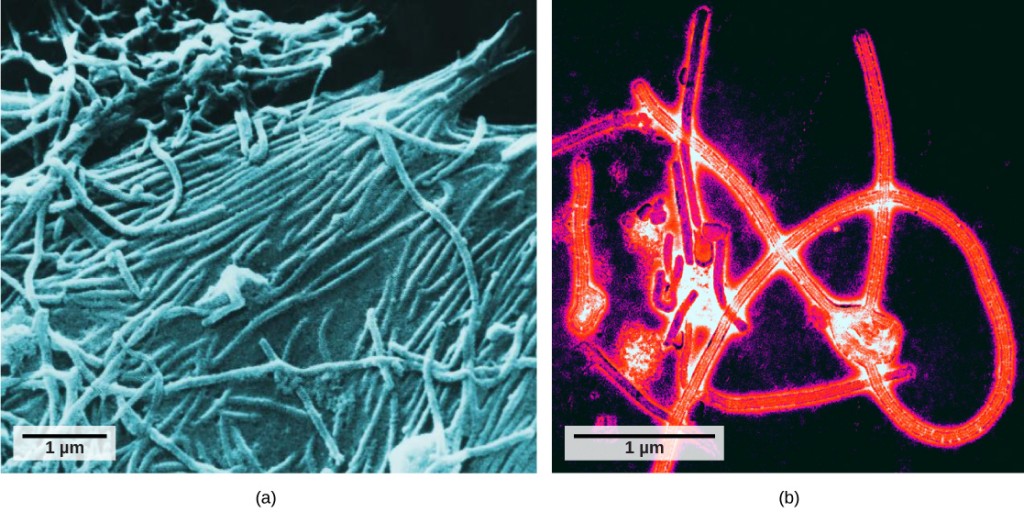
vasta elektronimikroskoopin kehityksen myötä 1940-luvulla tutkijat saivat ensimmäisen hyvän kuvan tupakan mosaiikkiviruksen rakenteesta (Kuva 12.2) ym. Virionien pintarakennetta voidaan tarkkailla sekä skannaus-että transmissioelektronimikroskopialla, kun taas viruksen sisäisiä rakenteita voidaan havaita vain transmissioelektronimikroskoopilla otetuista kuvista (Kuva 12.4).
tämän tekniikan käyttö on mahdollistanut monien virusten löytämisen kaikenlaisista elävistä organismeista. Ne ryhmiteltiin aluksi jaetun morfologian mukaan eli niiden koon, muodon ja erottavien rakenteiden mukaan. Myöhemmin virusten ryhmät luokiteltiin niiden sisältämän nukleiinihapon, DNA: n tai RNA: n tyypin mukaan ja sen mukaan, oliko niiden nukleiinihappo yksi – vai kaksijuosteinen. Viime aikoina viruksen replikaatiosyklien molekyylianalyysi on tarkentanut niiden luokittelua.
virioni koostuu nukleiinihappoytimestä, uloimmasta proteiinikuoresta ja joskus uloimmasta kuoresta, joka koostuu isäntäsolusta johdetuista proteiini-ja fosfolipidikalvoista. Näkyvin ero virusperheiden jäsenten välillä on niiden morfologia, joka on varsin monipuolinen. Mielenkiintoinen piirre viruksen monimutkaisuus on, että monimutkaisuus isäntä ei korreloi monimutkaisuus virionin. Joitakin monimutkaisimpia virionrakenteita havaitaan bakteriofageissa, viruksissa, jotka tartuttavat yksinkertaisimpia eläviä organismeja, bakteereja.
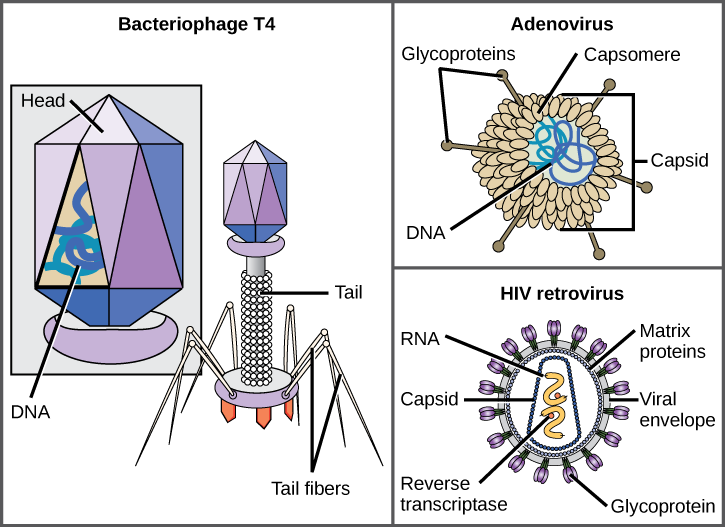
viruksia on monenmuotoisia ja-kokoisia, mutta ne ovat yhdenmukaisia ja erillisiä kunkin virusperheen osalta (Kuva 12.5). Kaikilla virioneilla on nukleiinihapon genomi, jota peittää suojaava proteiinikerros eli kapsidi. Kapsidi koostuu kapsomeereiksi kutsutuista proteiinialayksiköistä. Jotkin viruskapsidit ovat yksinkertaisia monitahokkaita ”sfäärejä”, kun taas toiset ovat rakenteeltaan melko monimutkaisia. Joidenkin virusten kapsidia ympäröivää ulompaa rakennetta kutsutaan viruksen kuoreksi. Kaikki virukset käyttävät jonkinlaista glykoproteiinia kiinnittyäkseen isäntäsoluihinsa solussa oleviin molekyyleihin, joita kutsutaan virusreseptoreiksi. Virus hyödyntää näitä solun pintamolekyylejä, joita solu käyttää johonkin muuhun tarkoitukseen, keinona tunnistaa ja infektoida tiettyjä solutyyppejä. Esimerkiksi tuhkarokkovirus käyttää ihmisellä solujen pinnalla olevaa glykoproteiinia, joka normaalisti toimii immuunireaktioissa ja mahdollisesti siittiön ja munasolun välisessä vuorovaikutuksessa hedelmöityksessä. Kiinnittyminen on vaatimus, että virukset tunkeutuvat myöhemmin solukalvoon, ruiskuttavat viruksen perimän ja viimeistelevät replikaationsa solun sisällä.
E. coli-bakteeria tartuttava T4-bakteriofagi on monimutkaisimpia tunnettuja virioneja; T4: ssä on proteiinihäntä, jota virus käyttää kiinnittyäkseen isäntäsoluun, ja päärakenne, jossa on sen DNA.
Adenovirus on ihmiselle hengityselinsairauksia aiheuttava epävelopaattinen eläinvirus, joka käyttää kapsomeereistaan ulkonevia proteiinipiikkejä kiinnittyäkseen isäntäsoluun. Nonenveloped viruksia ovat myös ne, jotka aiheuttavat polio (poliovirus), syylät (papilloomavirus), ja hepatiitti A (hepatiitti A-virus). Vieroittamattomat virukset ovat yleensä vankempia ja selviytyvät todennäköisemmin ankarissa olosuhteissa, kuten suolistossa.
Vaipalliset virionit kuten HIV (ihmisen immuunikatovirus), aidsin aiheuttaja (hankittu immuunivajausoireyhtymä), koostuvat nukleiinihaposta (HIV: n tapauksessa RNA) ja kapsidiproteiineista, joita ympäröi fosfolipidikerros ja siihen liittyvät proteiinit (Kuva 12.5). Vesirokko, influenssa ja sikotauti ovat esimerkkejä virusten aiheuttamista taudeista kuorineen. Kuoren haurauden vuoksi nonenveloped-virukset kestävät paremmin lämpötilan, pH: n ja joidenkin desinfiointiaineiden muutoksia kuin vaipalliset virukset.
kaiken kaikkiaan virionin muoto ja kuoren esiintyminen tai puuttuminen kertovat vain vähän siitä, mitä tauteja virukset voivat aiheuttaa tai mitä lajeja ne voivat tartuttaa, mutta on silti hyödyllinen keino aloittaa virusluokitus.
Mikä seuraavista viruksen rakennetta koskevista väitteistä on totta?
a) kaikki virukset koteloituvat viruksen kalvoon.
B) kapsomeeri koostuu pienistä proteiinialayksiköistä, joita kutsutaan kapsideiksi.
c) DNA on kaikkien virusten geneettinen materiaali.
d) glykoproteiinit auttavat virusta kiinnittymään isäntäsoluun.
<!- D – >
toisin kuin kaikki elävät organismit, jotka käyttävät DNA: ta geneettisenä materiaalinaan, virukset voivat käyttää joko DNA: ta tai RNA: ta omanaan. Viruksen ydin sisältää viruksen perimän eli koko geneettisen sisällön. Viruksen genomit ovat yleensä pieniä verrattuna bakteereihin tai eukaryootteihin, ja sisältävät vain niitä geenejä, jotka koodaavat proteiineja, joita virus ei voi saada isäntäsolusta. Tämä geneettinen materiaali voi olla yksijuosteinen tai kaksijuosteinen. Se voi olla myös lineaarinen tai pyöreä. Vaikka useimmat virukset sisältävät yhden nukleiinihapon segmentin, toisilla on genomeja, jotka koostuvat useista segmenteistä.
DNA-viruksilla on DNA-ydin. Viruksen DNA ohjaa isäntäsolun replikaatioproteiineja syntetisoimaan uusia kopioita viruksen perimästä ja litteroimaan ja kääntämään kyseisen perimän virusproteiineiksi. DNA-virukset aiheuttavat ihmisille sairauksia, kuten vesirokkoa, hepatiitti B: tä ja joitakin sukupuolitauteja, kuten herpes-ja genitaalialueen visvasyyliä.
RNA-virukset sisältävät vain RNA: ta ytimissään. Replikoidakseen genomejaan isäntäsolussa RNA-virusten genomit koodaavat entsyymejä, joita ei ole isäntäsoluissa. RNA-polymeraasientsyymit eivät ole yhtä stabiileja kuin DNA-polymeraasit ja tekevät usein virheitä transkription aikana. Tästä syystä RNA-viruksissa tapahtuu mutaatioita eli nukleotidisekvenssin muutoksia useammin kuin DNA-viruksissa. Tämä johtaa nopeampaan evoluutioon ja muutokseen RNA-viruksissa. Esimerkiksi se, että influenssa on RNA-virus, on yksi syy siihen, että uusi influenssarokote tarvitaan joka vuosi. RNA-virusten aiheuttamia ihmisten sairauksia ovat muun muassa hepatiitti C, tuhkarokko ja vesikauhu.
virukset voidaan nähdä velvoittavina solunsisäisinä loisina. Viruksen täytyy kiinnittyä elävään soluun, se täytyy viedä sen sisälle, valmistaa sen proteiinit ja kopioida sen genomi sekä löytää keino paeta solusta, jotta virus voi tartuttaa muita soluja ja lopulta muita yksilöitä. Virukset voivat tartuttaa vain tiettyjä isäntälajeja ja vain tiettyjä kyseisen isännän soluja. Tämän spesifisyyden molekyyli perustuu siihen, että isäntäsolun pinnalta on löydyttävä tietty pintamolekyyli eli virusreseptori, jotta virus kiinnittyy. Myös eri solutyypeissä havaitut metaboliset erot, jotka perustuvat erilaiseen geeniekspressioon, ovat todennäköinen tekijä, jossa virus voi monistaa soluja. Solun täytyy valmistaa viruksen tarvitsemia aineita, kuten entsyymejä, joille viruksen perimässä itsessään ei ole geenejä, tai virus ei pysty monistumaan kyseisen solun avulla.
virustartuntojen vaiheet
viruksen on ”vallattava” solu replikoituakseen. Viruksen replikaatiosykli voi aiheuttaa isäntäsolussa dramaattisia biokemiallisia ja rakenteellisia muutoksia, jotka voivat aiheuttaa soluvaurioita. Nämä muutokset, joita kutsutaan sytopaattisiksi vaikutuksiksi, voivat muuttaa solun toimintoja tai jopa tuhota solun. Jotkut tartunnan saaneet solut, kuten flunssaviruksen (rinoviruksen) tartuttamat, kuolevat lyysin (puhkeaminen) tai apoptoosin (ohjelmoitu solukuolema tai ”solu itsemurha”) kautta vapauttaen kaikki jälkeläiset virionit kerralla. Virustautien oireet johtuvat immuunivasteesta virukselle, joka yrittää hallita ja poistaa viruksen elimistöstä, sekä viruksen aiheuttamista soluvaurioista. Monet eläinvirukset, kuten HIV (human immunodeficiency virus), lähtevät immuunijärjestelmän infektoituneista soluista orastavalla prosessilla, jossa viriot poistuvat solusta yksitellen. Orastavan prosessin aikana solu ei käy läpi lyysiä eikä kuole heti. HI-viruksen tartuttamien solujen vaurioituminen voi kuitenkin tehdä solujen toimimisen immuniteetin välittäjinä mahdottomaksi, vaikka solut pysyisivätkin elossa jonkin aikaa. Tuottavimmat virusinfektiot noudattavat samanlaisia vaiheita viruksen replikaatiosyklin aikana: kiinnitys, tunkeutuminen, päällystäminen, replikointi, kokoonpano ja vapauttaminen.
virus kiinnittyy tiettyyn reseptoripaikkaan isäntäsolukalvolla kapsidin kiinnitysproteiinien tai sen kuoreen upotettujen proteiinien välityksellä. Kiinnittyminen on spesifistä, ja tyypillisesti virus kiinnittyy vain yhden tai muutaman lajin soluihin ja vain tiettyihin solutyyppeihin näiden lajien sisällä sopivilla reseptoreilla.
Concept in Action

katso videolta visuaalinen selitys siitä, miten HIV ja influenssa iskevät kehoon.
toisin kuin eläinvirukset, bakteriofagien nukleiinihappo ruiskutetaan isäntäsoluun alastomana, jolloin kapsidi jää solun ulkopuolelle. Kasvi-ja eläinvirukset voivat päästä soluihinsa endosytoosin kautta, jossa solukalvo ympäröi ja nielaisee koko viruksen. Osa vaipallisista viruksista pääsee soluun, kun viruskuori fuusioituu suoraan solukalvon kanssa. Solun sisälle päästyään viruksen kapsidi hajoaa ja viruksen nukleiinihappo vapautuu, joka sitten tulee saataville replikaatiota ja transkriptiota varten.
replikaatiomekanismi riippuu viruksen genomista. DNA-virukset käyttävät yleensä isäntäsolun proteiineja ja entsyymejä lisä-DNA: n valmistamiseen, jonka avulla genomi kopioidaan tai transkriboidaan lähetti-RNA: han (mRNA), jota sitten käytetään proteiinisynteesissä. RNA-virukset, kuten influenssavirus, käyttävät yleensä RNA-ydintä mallina virusten genomisten RNA: n ja mRNA: n synteesiin. Viruksen mRNA muuntuu virusentsyymeiksi ja kapsidiproteiineiksi uusien virionien kokoamiseksi (Kuva 12.6). Toki tähän kuvioon on poikkeuksia. Jos isäntäsolu ei tarjoa viruksen replikaatioon tarvittavia entsyymejä, viruksen geenit toimittavat tiedot puuttuvien proteiinien suorasynteesiin. Retroviruksilla, kuten HIV: llä, on RNA-genomi, joka on käännettävä transkriptioksi DNA: n valmistamiseksi, joka sitten työnnetään isännän DNA: han. RNA: n muuntamiseksi DNA: ksi retrovirukset sisältävät geenejä, jotka koodaavat viruspesifistä entsyymiä käänteiskopioijaentsyymiä, joka transkriboi RNA-mallin DNA: han. Se, että HIV tuottaa joitakin omia entsyymejä, joita ei löydy isännästä, on antanut tutkijoille mahdollisuuden kehittää lääkkeitä, jotka estävät näitä entsyymejä. Nämä lääkkeet, mukaan lukien käänteiskopioijaentsyymin estäjä AZT, estävät HIV: n replikaatiota vähentämällä entsyymin toimintaa vaikuttamatta isännän aineenvaihduntaan.
viruksen replikaation viimeinen vaihe on uusien virionien vapautuminen isäntäorganismiin, jossa ne pystyvät infektoimaan viereisiä soluja ja toistamaan replikaatiosyklin. Osa viruksista vapautuu isäntäsolun kuollessa ja toiset virukset voivat lähteä infektoituneista soluista orastamalla kalvon läpi tappamatta solua suoraan.
influenssavirus on pakattu viruskuoreen, joka fuusioituu plasmakalvon kanssa. Näin virus voi poistua isäntäsolusta tappamatta sitä. Mitä hyötyä virus saa pitämällä isäntäsolun elossa?
<!- Isäntäsolu voi jatkaa uusien virushiukkasten tuottamista.- >
Concept in Action

Napsauta tämän virusten opetusohjelman läpi tunnistaaksesi rakenteita, siirtymistapoja, replikaatiota ja paljon muuta.