”Kuinka kauan solusyklin eri vaiheet kestävät?
kuinka kauan solusyklin eri vaiheet kestävät?
lukutila
replikaatio on yksi elävän aineen tunnusmerkeistä. Joukko prosesseja kutsutaan solusyklin jotka toteutetaan yksi solu tulee kaksi on ollut hallitseva tutkimusaihe molekyyli aikakauden sovelluksia, jotka ulottuvat pitkälle myös tutkimukseen sairauksia, kuten syöpä, joka on joskus luonnehditaan sairaus solusyklin mennyt pieleen. Solusyklit ovat mielenkiintoisia sekä sen vuoksi, miten ne ovat samanlaisia solutyypistä toiseen, että sen vuoksi, miten ne ovat erilaisia. Jotta aihe helpottuisi, otamme huomioon useiden eri eliöiden solusyklit, mukaan lukien prokaryootin mallin, nisäkässoluille kudosviljelmässä ja hedelmäkärpäsen alkionkehityksen aikana. Kysymme erityisesti, mitkä ovat ne yksittäiset vaiheet, joihin ryhdytään, jotta yksi solu jakaantuisi kahteen ja kuinka kauan nämä vaiheet kestävät?
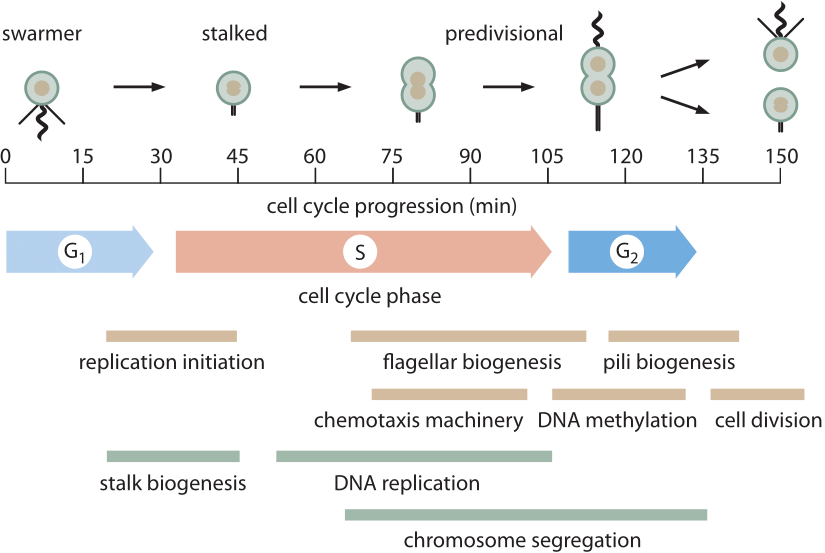
kuva 1: Caulobacterin 150 min solusykli on esitetty korostaen joitakin keskeisiä morfologisia ja metabolisia tapahtumia, jotka tapahtuvat solunjakautumisen aikana. M-vaihetta ei ole merkitty, koska Caulobacterissa ei ole varsinaista mitoottista laitetta, joka koottaisiin kuten eukaryooteissa. Suuri osa caulobacterin (ja muiden bakteerien) kromosomien erottelusta tapahtuu samanaikaisesti DNA: n replikaation kanssa. Kromosomien erottelun ja erityisesti kahden kehäkromosomien dekatenaation viimeiset vaiheet tapahtuvat G2-vaiheen aikana. (Mukautettu M. T. Laub et al., Tiede 290:2144, 2000.)
luultavasti parhaiten tunnettu prokaryoottisolusykli on malliorganismilla Caulobacter crescentus. Yksi tämän bakteerin houkuttelevista piirteistä on se, että siinä on epäsymmetrinen solunjakautuminen, jonka avulla tutkijat voivat sitoa toisen jälkeläisistä mikroskoopin suojuslipukkeeseen, kun taas toinen tytär ajelehtii pois mahdollistaen lisätutkimuksen ilman esteitä. Tämä on johtanut huolelliseen kuvaukseen ≈150 minuutin solusykli (bnid 104921), kuten kuvassa 1 esitetään. Solusyklin pääkomponentit ovat G1 (ensimmäinen kasvuvaihe, ≈30 min, bnid 104922), jossa solun koon täytyy kasvaa ainakin jonkin verran, s-vaihe (synteesi, ≈80 min, bnid 104923), jossa DNA replikoituu, ja G2 (toinen kasvuvaihe, ≈25 min, bnid 104924), jossa kromosomien eriytyminen etenee johtaen solunjakautumiseen (viimeinen vaihe kestää ≈15 min). Caulobacter crescentus on kiinnostava esimerkki tavasta, jolla tietyt organismit ylennetään ”malliorganismiksi”, koska niillä on jokin erityinen ominaisuus, joka tekee niistä erityisen otollisia kiinnostuskysymykselle. Tässä tapauksessa solusyklin eteneminen kulkee käsi kädessä erilaistumisprosessin kanssa, joka antaa helposti visualisoitavia tunnistettavia vaiheita, mikä tekee niistä parempia solusyklin biologeille kuin vaikkapa mallibakteeri E. coli.
nisäkässolujen käyttäytyminen kudosviljelmissä on toiminut pohjana paljolti sille, mitä tiedämme korkeammissa eukaryooteissa solusyklistä. Eukaryoottinen solusykli voidaan jakaa karkeasti kahteen vaiheeseen, interfaasiin, siihen solusyklin osaan, kun solun materiaaleja monistetaan, ja mitoosiin, fysikaalisten prosessien joukkoon, jotka osallistuvat kromosomien erotteluun ja sitä seuraavaan solunjakautumiseen. Solusyklin prosessien nopeudet rakentuvat enimmäkseen monista molekyylitapahtumista, kuten DNA: n ja sytoskeletaalisten filamenttien polymeroitumisesta, joiden nopeuksia olemme jo tarkastelleet. Hela-kennon tyypillisestä 20 tunnin solusykliajasta lähes puolet on omistettu G1: lle (BNID 108483) ja lähes toinen puoli on S-vaihe (BNID 108485), kun taas G2 ja M ovat paljon nopeampia noin 2-3 tunnin ja 1 tunnin kohdalla (BNID 109225, 109226). Kestoltaan vaihtelevin vaihe on G1. Epäedullisemmissa kasvuolosuhteissa, kun solusyklin kesto kasvaa, tämä on vaihe, joka vaikuttaa eniten, luultavasti johtuen ajasta, joka kestää jonkin säädellyn koon tarkistuspisteen saavuttamiseen. Vaikka erityyppiset todisteet viittaavat tällaisen tarkastuspisteen olemassaoloon, se tunnetaan nykyisin hyvin huonosti. Historiallisesti solusyklin vaiheet on yleensä päätelty kiinteiden solujen avulla, mutta viime aikoina geneettisesti koodatut biosensorit, jotka muuttavat lokalisaatiota solusyklin eri vaiheissa, ovat mahdollistaneet elävien solujen ajallisen tiedon solusyklin etenemisestä ja pysäyttämisestä.
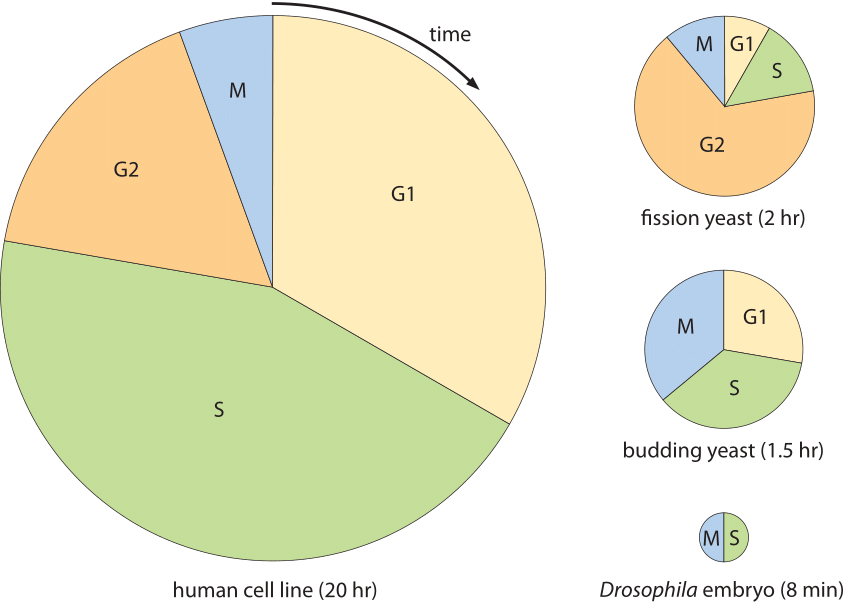
kuva 2: solusyklin ajat eri solutyypeille. Jokainen piirakaavio näyttää solusyklin murto-osan, joka on omistettu kullekin solusyklin primaarivaiheelle. Kunkin kaavion pinta-ala on verrannollinen kokonaissolusyklin kestoon. Solusyklin kestot heijastavat minimaalisia kaksinkertaistumisaikoja ihanteellisissa olosuhteissa. (Sovitettu David Morganin kappaleesta ”The Cell Cycle – Principles of Control”.)
miten solusyklin pituus vertautuu aikaan, joka Solulta kuluu uuden genominsa syntetisoimiseen? Eukaryooteilla on perimän pituuden ja kaksinkertaistumisajan välinen irrotus johtuen useiden DNA: n replikaation aloituspaikkojen käytöstä. Nisäkässoluissa on havaittu, että monissa kudoksissa, joiden kokonaissykliaika vaihtelee suuresti, DNA: n replikaation s-vaiheen kesto on huomattavan vakio. Hiiren kudoksissa, kuten paksusuolessa tai kielessä, S-vaihe vaihteli pienellä alueella 6,9-7,5 tunnissa (BNID 111491). Jopa verrattaessa useita epiteelikudoksia ihmisen, rotan, hiiren ja hamsterin välillä, S-vaihe oli 6-8 tuntia (BNID 107375). Nämä mittaukset tehtiin 1960-luvulla tekemällä eräänlainen pulssijahtikoe radioaktiivisesti merkityllä nukleotiditymidiinillä. Lyhyen pulssin aikana radioaktiivinen yhdiste imeytyi vain S-vaiheen solujen genomiin. Mittaamalla kesto ulkonäkö ja sitten katoaminen merkitty solujen m-vaiheessa voidaan päätellä, kuinka kauan S-vaihe kesti se, että kesto s-vaihe on suhteellisen vakio tällaisissa soluissa käytetään tähän päivään arvioida kesto solusyklin alkaen tietoa vain murto solujen tietyn tilannekuvan ajan, jotka ovat S-vaiheessa. Esimerkiksi, jos kolmasosa soluista nähdään s-vaiheessa, joka kestää noin 7 tuntia, solusyklin ajan on päätelty olevan noin 7 tuntia / (1/3) ≈20 tuntia. Nykyään tällaiset mittaukset tehdään useimmiten käyttämällä BrdU: ta s-vaiheen merkkinä. Emme tiedä tyydyttävää selitystä tämän suhteellisen jatkuvan replikaatioajan synnylle ja sille, miten se liittyy DNA-polymeraasin nopeuteen ja replikaation aloituspaikkojen tiheyteen genomin varrella.
Solusyklien moninaisuus on esitetty kuvassa 2 ja siinä kuvataan useita malliorganismeja sekä niiden solusyklien eri vaiheiden kestoja ja sijoittumista. Ääriesimerkki esiintyy hedelmäkärpäsen Drosophila melanogasterin alkionkehitysprosessissa. Tässä tapauksessa tilanne poikkeaa tavanomaisista solunjakautumisista, sillä uusien sytoplasmamateriaalien syntetisoinnin sijaan massa säilyy olennaisesti geneettisen materiaalin replikaatiota lukuun ottamatta. Tämä tapahtuu hyvin synkronisesti noin 10 sukupolven ajan, ja alkion tuhansien solujen replikaatiosykli, esimerkiksi syklien 10 ja 11 välillä, tapahtuu noin 8 minuutissa, kuten kuvassa 2 (BNID 103004, 103005, 110370). Tämä on nopeampi kuin yhdenkään bakteerin replikaatioajat, vaikka genomi on ≈120 miljoonaa bp pitkä (BNID 100199). Näyttävä esimerkki solujen kyvystä mukauttaa aikadynamiikkaansa.




