Solubiologia 07: Mikrotubulukset ja Solunjakautuminen
nämä ovat Harvard Extension ’ s Cell Biology course-luennon 7 muistiinpanoja.
Luento 6 esitteli Mikrotubulukset, ja tässä luennossa käsitellään niiden roolia solunjakautumisessa. Tässä esittelyvideo:
yleiskatsaus solusykliin
solusykliä – solujen jakautumisen ja replikaation prosessia – säätelee sarja biokemiallisia kytkimiä, joita kutsutaan solusyklin ohjausjärjestelmäksi.
solusykli on jaettu vaiheisiin, jotka jakautuvat vaiheisiin – ihmiset tarkoittavat ”4 vaihetta”, mutta sitten niitä on todellisuudessa 5, ja ihmiset käyttävät myös muita sanoja ryhmitelläkseen nämä vaiheet yhteen, ja toisin sanoen sanoja alittaakseen ne. Olen tehnyt parhaani tiivistää suhde näiden termien alla olevassa taulukossa. (muokattu/laajennettu Wikipediasta):
| MOST general grouping | the supposed ”4 phases” | subphases |
|---|---|---|
| non-dividing | Gap 0 (G0) | |
| interphase | Gap 1 (G1) | G1a R G1b |
| Synthesis (S) | ||
| Gap 2 (G2) | ||
| Mitosis | Mitosis (M) | prophase prometaphase metaphase anaphase telophase cytokinesis |
The kunkin vaiheen sisältö on kauniisti tiivistetty tähän Kelvinsongin erinomaiseen Wikimedia Commons-kuvaan:

nopeimmin jakautuvat ihmissolut voivat suorittaa solusyklin noin 24 tunnissa (G1: 9h, S: 10h, G2: 4h, M: 30 min). Hiiva voi päättää syklin 30 minuutissa, ja nopeimmin jakautuvissa Drosophila-soluissa kestää jopa 8 minuuttia.
tämän prosessin Pääohjaimiin kuuluvat sykliinit, jotka säätelevät sykliiniriippuvaista kinaasia eli CDK: ta. Muista, että kinaasit ovat proteiineja, jotka fosforyloivat muita proteiineja. CDK: n kohteiden fosforylointi mahdollistaa mitoosin etenemisen. Tarkemmin sanottuna kypsymistä edistävä tekijä eli MPF on sykliini B: n ja CDK: n muodostama obligaatti heterodimeerinen kompleksi, joka tekee fosforylointitoimintansa vain molempien proteiinien ollessa läsnä.
mikrotubulusten rooli
Mikrotubulukset ovat kriittisiä koko solusyklin ajan – ne organisoivat solukomponentit ja jakavat ne kahtia. Tässä on joukko videoita solusyklin, jotka korostavat mikrotubulusten roolia:
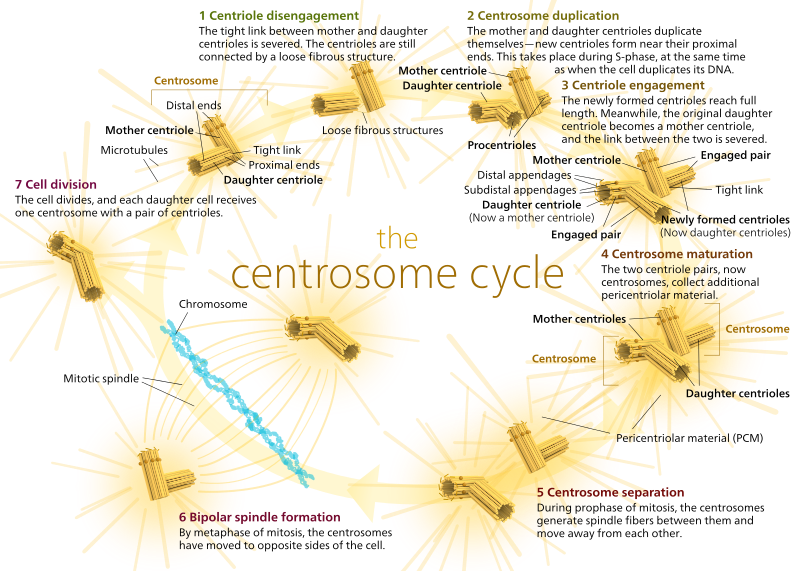
eläimillä leposoluissa ja jopa interfaasin soluissa on yleensä vain yksi mtoc eli centrosomi, joka toimii solun kaikkien mikrotubulusten keskuskeskuksena. Centrosome koostuu kahdesta centriolista kuten alla (kiitos jälleen Kelvinsongille):

kaksi centriolia irtaantuvat toisistaan ja monistuvat S-vaiheen aikana ja sitten erkanevat muodostaen solun vastakkaisia”napoja”m-vaiheen aikana niin, että nyt on kaksi Mtocia, joista kummastakin tulee lopulta uuden solun ainoa MTOC (toinen boss Kelvinsong-kuva):
mitoosin aikana on siis solun kaksi”napaa”, joissa kummassakin on Mikrotubulukset ankkuroituina (-) päähän ja niiden (+) päät päällekkäisinä osoittaen solun keskelle, kuten tästä näkyy (Wikimedia Commons image by Lordjuppiter):
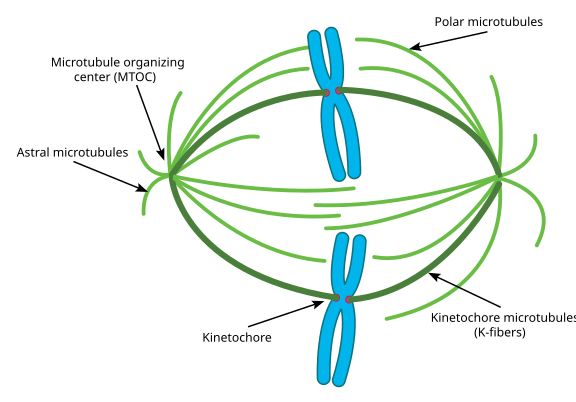
tuota kokonaisuutta kutsutaan karakoneeksi, ja aluetta, jossa kaksi MTOCs: n mikrotubulusta limittyvät, kutsutaan interdigitaation vyöhykkeeksi. Joskus kuulee jokaisen MTOC: n ja sen merisiilimäisen mikrotubulusten joukon, jota kutsutaan mitoottiseksi asteriksi.”
mikrotubulusten sanotaan tässä vaiheessa jakautuvan kolmeen luokkaan:
- astraaliset Mikrotubulukset osoittavat ulospäin, kohti solukuorta, jotta koko karakoneisto kiinnittyy solunjakautumisakselia pitkin.
- Kinetokorin Mikrotubulukset kiinnittyvät kromatidien kinetokoriin.
- polaariset Mikrotubulukset, jotka ovat suuntautuneet yhdensuuntaisesti mutta vastakkaisiin suuntiin, ovat ratkaisevia painettaessa karakonetta erilleen mitoosin aikana. (Itse asiassa polaariset Mikrotubulukset ovat myös läsnä aikaisemmin ja auttavat työntämään sentrosomit erilleen profaasin aikana).
Jos haluat mieluummin valokuvia kuin diagrammeja, koko karakoneisto näyttää tältä: kromatidit sinisinä, Mikrotubulukset vihreinä ja kinetokorit punaisina pisteinä:

Mikrotubulukset muuttuvat paljon dynaamisemmiksi mitoosin aikana: enemmän gammatubuliinia edistää helpompaa nukleaatiota, mutta XMAP215, mikrotubulusstabilaattori, fosforyloituu ja siten inaktivoituu mitoosin aikana, jolloin Kinesiini-13 voi vapaasti katastrofoida Mikrotubulukset. Omaisuuksia tehdään ja menetetään nopeasti. Mikrotubuluksen puoliintumisaika mitoosin aikana on noin 15 minuuttia, kun se interfaasin aikana on 30 minuuttia. Ihmiset tutkivat mikrotubulusdynamiikkaa FRAP: lla: lisää fluoresoiva mikrotubuli, valkaise se ja katso kuinka nopeasti uudelleenkokoontuminen tapahtuu sen perusteella, kuinka pian fluoresenssi ilmestyy uudelleen. + Vinkeillä on myös suuri merkitys mikrotubulusten auttamisessa ja kokoamisessa.
Kinesiini-5: llä on kaksi napapäätä, jotka sitoutuvat vastakkaisiin mikrotubuluksiin ja yrittävät kävellä kummankin (+) päätä kohti. Tämä työntää kaksi mikrotubulusta erilleen ja antaa liikkeellepanevan voiman MTOCs: n erottamiseen.
Sentromeerisessa DNA: ssa on vähän informaatiota entropiaa ja erityisiä histoneja, jotka eroavat muista kromatiineista. Centromeerit ovat yksi osa perimää, jota ei juuri koskaan opi seuraavan sukupolven sekvensoinnissa, edes todella korkealla syvyydellä. Tämä johtuu siitä, että sentromeerit palvelevat eri tarkoitusta kuin suuri osa muusta genomista: sekvenssi siellä on suotuisa vuorovaikutukselle sentromeeristen proteiinien ja kinetokorin kiinnittymisen kanssa. Koossiinit ovat proteiineja, jotka pitävät kaksi sisarkromatidia yhdessä. Kinetokoriproteiineissa on kaksi kerrosta, sisempi kinetokori ja ulompi kinetokori.
Prometafaasin aikana kromosomit liikkuvat edestakaisin. Kinesiinit ankkuroivat kromosomit kinetokorin mikrotubuluksiin kärjen ulkopuolelle, missä Kinesiini-13 depolymeroi Mikrotubulukset, apunaan käytettävissä olevien tubuliinidimeerien puute. Yhdistelmä motorisia proteiineja, mikrotubulus vuorovaikutuksessa proteiineja ja juoksumatolla palvelee siirtää kromosomit. Samaan aikaan dynein ja dynactin – motoriset proteiinit, jotka kävelevät kohti (-) päätettä, toimivat astraalisissa mikrotubuluksissa vetäen Mtoceja kohti solun kehää. Metafaasissa kromatidit tulevat kohdistumaan ”metafaasilevyä” pitkin.
tämän prosessin aikana ydinkuori liukenee, joten ydinvoiman tuonnista tulee merkityksetöntä. Ran-GEF paikantaa lähellä kromosomeja ja tuottaa suuria pitoisuuksia Ran-GTP: tä, joka tuottaa energiaa joihinkin välttämättömiin prosesseihin (?).
soluilla on jokin mekanismi mikrotubulusten jännityksen havaitsemiseksi, joka osoittaa niiden kiinnityskromatidit ennen kuin mitoosi voi edetä. Varmista, että jokainen kromatid on oikein ankkuroitu on tärkeää välttää aneuploidy.
muuten muillakin sytoskeletaaleilla on mikrotubulusten lisäksi keskeinen rooli solusyklissä. Sytokineesissä aktiini muodostaa supistuvan renkaan ja myosiini II: n motoristen proteiinien avulla cinches solun kahdeksi.
Malliorganismien merkitys
solusyklin säätelyprosessien keksiminen perustui vahvasti joihinkin suosittujen malliorganismien siisteihin piirteisiin.
Saccharomyces cerevisiae (orastava hiiva) ja Schizosaccharomyces pombe (fissiohiiva) voivat esiintyä haploideina tai diploideina. Se on tärkeää, koska haploidivaiheessa yksi mutaatio voi tyrmätä geenin – sinun ei tarvitse osua molempiin alleeleihin. Hiivassa monet mutaatiot, erityisesti Cdc__ (cell division control)-geeneissä, ovat lämpötilariippuvaisia, joissa missense-mutaation omaava proteiini voi vielä toimia oikein ”sallivissa” lämpötiloissa, mutta menettää alkuperäisen tehtävänsä ”ei-sallivissa” lämpötiloissa. Tämä mahdollistaa knockout-fenotyypin tutkimisen (ei-sallivassa lämpötilassa) samalla kun sillä on se mukavuus, että se pystyy helposti levittämään eliöitä (sallivassa lämpötilassa). Koko S. cerevisiae-genomi on saatavilla plasmidikirjastoina, mikä mahdollistaa seulonnan, jolle plasmidi pelastaa tietyn mutantin fenotyypin. Näin löydettiin monia solusykliä sääteleviä geenejä.
In S. cerivisiae, orastava on osa vaihetta G1, ja kun tytär solu saavuttaa tietyn koon, hetkellä kutsutaan ”alkaa”, kaksi ovat sitoutuneet syöttämään S ja lopulta loppuun solusyklin. Nisäkässoluilla on oma sitoutumispisteensä, jota kutsutaan G1: ssä rajoituspisteeksi tai R: ksi, joka on analoginen aloittamaan.
Lämpötilaherkät Cdc28-mutantit eivät nupahda epäpermissiivisessä lämpötilassa. Cdc28-geeni koodaa hiivan sykliiniriippuvaisen kinaasin (CDK) homologeja, jotka sykliinillä kompleksoituna voivat fosforyloida muita proteiineja säätelemään niiden osallistumista solusyklin vaiheisiin. Lämpötilaherkät mutantit ei-läpäisevässä lämpötilassa juuttuvat kykenemättömiksi nousemaan ja astumaan S-vaiheeseen. Sen sijaan ne käyttäytyvät kuin villityyppiset solut, joilta on riistetty ravinteet: ne kasvavat tarpeeksi isoiksi läpäistäkseen alun, mutta eivät sitten Jatka.
Xenopus (sammakkolaji) osoittautui kriittiseksi solusyklin ymmärtämisessä, koska sen lisääntymiseen liittyy hyvin suuri määrä soluja (ts. riittävästi lähtöainetta länkkärit yms.), jotka ovat täydellisesti synkronoituja (eli kaikki ovat samassa solusyklin vaiheessa samalla hetkellä. (Vertaa esimerkiksi hiivaan, jossa kaikki solut eivät ole samassa vaiheessa samaan aikaan). Myös itse muna on suuri ja helppo työstää, ja hedelmöitystä seuraa useita solusyklejä. Sammakoissa munat alkavat jakautua meioottisesti, mutta pysähtyvät sitten G2-vaiheessa 8 kuukaudeksi, kun ne kasvavat kooltaan ja varastoivat asioita, joita tarvitaan kasvuun hedelmöittymisen jälkeen.
Välifilamentit
mikrofilamenttien ja mikrotubulusten lisäksi eukaryoottisoluissa on myös joukko ”muita” sytoskeletaalisia proteiineja, joita kutsutaan välifilamenteiksi (IFS). Vaikka IFs on monimuotoisempi kuin mikrofilamentit ja Mikrotubulukset, se ei ole vain ”minkä tahansa muun hehkulangan” yleisnimitys, vaan se on ryhmä toisiinsa liittyviä proteiineja. Ne ulottuvat yleensä sytoplasman ja sisemmän ydinkuoren läpi, ovat nonpolaarisia eikä niihin liity motorisia proteiineja. Niillä on suuri vetolujuus ja ne ovat hyvin vakaita, hidas vaihtokurssi eikä paljon hajoamista, vaikka fosforylaatio voi edistää niiden purkamista. Tässä muutamia suosittuja esimerkkejä:
- keratiineja on epiteelisoluissa, mesodermin soluissa ja neuroneissa. Ne antavat voimaa ja tulevat happamissa ja perusmuodoissa. Kukin voi muodostaa oman juosteensa, mutta useimmat if: t koostuvat kahdesta säikeestä – yhdestä emäksisestä ja yhdestä hapokkaasta, tavallaan toistensa ympärille kiertyneestä säikeestä. Hiukset ja kynnet on valmistettu ”kova” keratiini runsaasti kysteiiniä disulfidi sidoksia, joka tarjoaa valtava vahvuus. Permanentit ja suoristus perustuvat disulfidisidosten vähentämiseen, hiusten muokkaamiseen ja sitten disulfidisidosten uudistamiseen. Ihossasi on myös ”pehmeää” keratiinia.
- desmiinejä, kuten vimentiiniä, on mesenkyymisoluissa (luu, kärtiledge ja rasva).
- Neurofilamentit ovat neuronaalisissa aksoneissa ja säätelevät niiden halkaisijaa, mikä puolestaan määrää aktiopotentiaalin etenemisnopeuden.
- Lamiinit ovat molemmat laajimmalle levinneitä, ja niiden uskotaan muistuttavan eniten kaikkien muiden if: ien fylogeneettistä kantamuotoa. Ne antavat rakenteellista tukea ydinkalvolle. Ne voivat auttaa avaruuden huokoskomplekseja ja myös organisoida DNA: ta.
lopuksi koostevideo: